Yawning is contagious. However, little
research has been done to elucidate the neuronal
representation of this phenomenon. Our study
objective was to test the hypothesis that the
human mirror neuron system (MNS) is activated by
visually perceived yawning. We used functional
magnetic resonance imaging to assess brain
activity during contagious yawning (CY).
Signal-dependent changes in blood oxygen levels
were compared when subjects viewed videotapes of
yawning faces as opposed to faces with a neutral
expression. In response to yawning, subjects
showed unilateral activation of their Brodmann's
area 9 (BA 9) portion of the right inferior
frontal gyrus, a region of the MNS. In this way,
two individuals could share physiological and
associated emotional states based on perceived
motor patterns. This is one component of empathy
(motor empathy) that underlies the development
of cognitive empathy. The BA 9 is reportedly
active in tasks requiring mentalizing abilities.
Our results emphasize the connection between the
MNS and higher cognitive empathic functions,
including mentalizing. We conclude that CY is
based on a functional substrate of empathy.
Introduction
Little research has been done to elucidate
an origin for the fascinating phenomenon of
contagious yawning (CY) (Provme 1986). In
contrast to spontaneous yawning, which is
considered evolutionarily old (Vischer 1959;
Sepulveda and Mangiamarchi 1995), CY is
phylogenetically and ontogenetically young, and
may not appear until the second year after birth
(Piaget 1951; Provme 1989; Anderson and Meno
2003). Whereas CY occurs in only a limited
number of animal species besides humans,
including chimpanzees (Anderson et al. 2004),
macaques (Paukner and Anderson 2006), baboons
(Palagi et al. 2009), and dogs (Joly-Mascheroni
et al. 2008), spontaneous yawning can be found
in almost all vertebrates. Why does CY require
such a high degree of evolutionary and
developmental specialization? CY is an
interaction between two individuals, with one
person experiencing and sharing the
physiological and emotional state of the other,
and a mechanism for synchronizing the state of a
group. This implicit link between two persons in
CY is considered an easily observable sign of
empathy (Lehmann 1979; Provine 2005; Senju 2010;
Arnott et al. 2009).
CY is impaired in children with autism
spectrum disorder (Senju et al. 2007; Senju et
al. 2009), patients with PTSD (Nietlisbach et
al. 2010), and those with schizophrenia (Haker
and Rössler 2009) or schizotypal
personality traits (Platek et al. 2003). All of
these conditions are accompanied by reduced
empathic abilities. Currently accepted concepts
of empathy state that contagion constitutes one
functional component of empathy-motor
empathy-and is mediated by brain areas involved
in the mirror neuron system (MNS) (Gallese 2007;
Preston and de Waal 2002; Leslie et al. 2004;
Blair 2005; Decety and Lamm 2006; Keysers and
Gazzola 2007; Uddin et al. 2007; Haker et al.
2010)
The MNS is a network of visuo-motor neurons
that was first discovered in a macaque in area
F5 of the pre-motor cortex (Rizzolatti et al.
1996). These neurons are active when a
particular action is performed or when the same
action, done by another individual, is observed.
Mirror neurons with similar properties have been
found in the posterior parietal cortex,
reciprocally connected with area F5 (Rizzolatti
et al. 2001). Experimental evidence suggests
that an analogous action observation-execution
matching system exists in humans. Studies using
electroencephalography, trans-cranial magnetic
stimulation, positron emission tomography, and
functional magnetic resonance imaging (fMRI)
have revealed a network composed of the pars
opercularis of the inferior frontal gyrus (IFG),
the anterior part of the inferior parietal
lobule (IPL), and the superior temporal sulcus
(STS) (Rizzolatti and Sinigaglia 2010).
Because one's own motor patterns can be
activated while observing an individual and
anticipating its effect from the same
perspective as the one who is acting, the mirror
mechanism generates the basis for shared
perception (Gallese 2003). In this way not only
simple motor actions but also emotional states
can be shared, as if by contagion, between human
beings (Can et al. 2003). By applying video
sequences, Platek et al. (2005) have found
bilateral activity in the posterior cingulate
and in the precuneus of individuals exposed to
yawning faces contrasted to laughing faces.
These regions belong to a medial fronto-parietal
network that mediates processes focused on
internal, mental, emotional, and experiential
characteristics of others or oneself (Lieberman
2006). Schürmann et al. (2005) have
reported that the right STS is activated when a
person is stimulated by a video-taped yawning
face but not one that is performing similar
non-yawning mouth movements. The STS is a region
of the externally oriented fronto-parietal
network, which is thought to represent the main
visual input to the MNS and to detect
specifically socially meaningful stimuli
(lacoboni 2005).
Our aim was to search for possible
activation of regions associated with the MNS,
as IFG (as a motor core of the human MNS), as
well as the IPL and STS (Rizzolatti and
Craighero 2004), during visual contagion by
yawning. This mechanism, as hypothesized by
Cooper et al. (2008), has been found in auditory
contagious yawning by Arnott et al. (2009) but,
according to our knowledge, has not yet been
verified in a visual paradigm.
To compare the effects of stimulations, we
used video sequences that depicted yawning faces
in contrast to faces showing minimal,
physiological, smooth-head, -mouth, and -gaze
movements by a person scanning the environment
without emotional mimic expression (i.e., a
non-contagious biological motion). We conducted
fMRI to monitor changes in blood oxygen
level-dependent (BOLD) signals. In contrast to
the above-mentioned study by Platek et al.
(2005), who contrasted a neutral condition
against two contagious conditions, yawning and
laughing, we considered our contrast to be more
specific to the contagious potential of the
yawning stimulus. Thus, we hypothesized that the
BOLD signal would increase in regions attributed
to the MNS when persons viewed yawning faces but
not faces with neutral expressions.
Discussion
We tested the hypothesis that the MNS is
activated when persons view CY. Changes in BOLD
signaling were investigated in the regions
attributed to the MNS while the subjects watched
video-taped yawning faces, neutralexpression
faces, and a baseline condition of static
scrambled face pictures. We found bilateral
activations in regions reported to be involved
in the human MNS and in face perception (i.e.,
inferior frontal and middle temporal gyms) in
both dynamic-stimulation conditions (neutral
expression faces and yawning faces) contrasted
to the static baseline (Rizzolatti and Craighero
2004; Talairach and Toumoux 1988; Puce et al.
1998). The response to faces with neutral
expressions included only minimal,
physiological, and smooth movements of the head,
mouth, and eyes. These are motions associated
with an individual who is quietly scanning the
environment and whose perception activates MNS
regions. This finding is in accord with that of
Nahab et al. (2009), who reported MNS activation
under both yawning and non-yawning (gape and
cough) conditions. The difference between our
conditions of yawning and neutral faces was
assumed to be the effect of contagiousness.
On the behavioral level, a contagion was
indicated in approximately 55 % of the
stimulations. This rate is comparable to that
described by Provine (1989) and Platek et al.
(2003), whose stimuli were rated within similar
settings. Because contagion is considered to be
primarily an automatic phenomenon, with a
conscious cognitive process being only a
secondary response, we based our evaluation on
all trials rather than just those where a
conscious feeling of contagion was indicated or
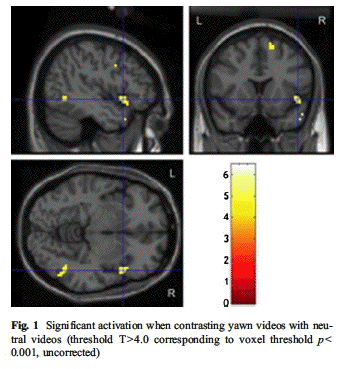
on a comparative analysis. When we contrasted
the two conditions (yawning vs. neutral), we
found only right-sided activation: besides
activation of the middle temporal gyrus, we
found specific activation in the BA 9 portion of
the right IFG and in the right superior frontal
gyms. The BA 9 is involved in higher social
cognitive functioning such as mentalizing
(Ohnishi et al. 2004). Thus, we concluded that
activation of this area in our contrast might
represent the effect of contagiousness, possibly
linking the MNS to higher cognitive functions
such as cognitive empathy (Haker et al. 2010).
This involvement of an area associated with
higher cognitive functions, which are not
developed at birth, may explain why CY is
ontogenetically seen only in later stages of a
person's development.
The right hemispheric dominance for
processes involving mentalizing is supported by
our results and is also in accordance with
results from neuropsychological studies on
hemispheric lesions (Siegal and Varley 2002). We
interpret the other specific activation in the
right superior frontal gyrus as representing the
suppression of the urge to yawn during the
experiment. Beauregard et al. (2001) reported
activation of this region during a task of
volitional inhibition of a comparable vegetative
reaction induced by visual stimulation, i.e.,
sexual arousal. By comparison, for motor tasks
such as finger movements, the temporo-parietal
junction and the anterior fronto-median cortex
have been identified as involved in inhibiting
imitation (Brass et al. 2009). However, these
movements do not elicit a vegetative urge such
as yawning or sexual stimuli. Therefore, other
mechanisms may be involved here.
The absence of MNS activation in CY has been
described in previous imaging studies by Platek
et al. (2005) and Schürmann et al. (2005).
This might be explained because those earlier
tests contrasted yawning with two other
potential MNS activators (Platek: laughing;
Schürmann: mouth movements similar to
yawning), as has already been discussed by
Arnott et al. (2009). The finding by the Platek
group of activation in the cortical midline
structures supports their "empathic modeling
hypothesis" of CY. This concept considers
contagious yawning to be "a primitive form of
empathic modeling that is subserved by
substrates that are precursors to a more
sophisticated and distributed system involved in
conscious self-processing", i.e., an element of
cognitive empathy (Platek et al. 2003). In line
with Platek, we consider our evidence for BA 9
activation during CY as a bottom-up input for
cognitive empathy and as a basis for such
higher-level aspects of cognitive empathy, e.g.,
conscious self-processing or the attribution of
mental states to other persons (Gallese 2007;
Haker et al. 2010). Schürmann et al. (2005)
have reported IFG activation in both stimulation
conditions when contrasted to a baseline.
Therefore, the IFG was no longer seen in the
contrast between those conditions.
Schürmann et al. explained this STS
activation when contrasting the two conditions
as evidence of an affinity in this region to
socially meaningful cues (in this case,
yawning). They conclude that "viewing another
person yawn seems to circumvent the essential
parts of the MNS, in line with the nature of
contagious yawns as automatically released
behavioral acts-rather than truly imitated motor
patterns". However, the behavioral act (i.e.,
the manifest yawn) did not occur during the
scanning in their study either, as participants
were instructed to avoid head movements. We
interpret the difference between their two
conditions as the potential of the true yawn
stimulus to elicit a highly stereotypical
vegetative reaction based on the activation of
the MNS, whereas the mere yawn-similar mouth
movements lead to a comparable MNS (IFG)
activation that lacks this potential. After the
scanning, their participants had to rate their
covert tendency to yawn during the scanning.
There, they indicated a greater tendency to yawn
during the yawn vs. the control condition.
However, their urge to imitate covertly the
other mouth movements in the control condition
was not reported. Based on the IFG activation in
the control condition, we assume that the
tendency to imitate those mouth movements was
also present in the control condition.
In addition to the results described here,
we must also address some limitations. One might
argue that the activation observed under our
test conditions might have been due to
participants observing mouth movements
associated with yawning, such reflecting mere
movement observation. However, a major function
of the MNS is to copy and extract the goal of
observed movements in order to behave
intuitively or automatically like the person
being observed. Thus, the associated activity
can be interpreted as yawning-related mirror
neuron activity because the contagious element
represents yawning-associated mouth movements.
With regard to the stimuli used here, we cannot
deny that the yawning videos were inherently
more interesting than the neutral videos. This
may have influenced the level of activity
observed during stimulation with yawning vs.
neutral videos. However, we did not find any
attention-specific differences in activation
patterns under those two conditions.
Our examination was further hindered because
of an essential methodological issue, for which
we had to ask that the subjects not perform
yawning motions in order to avoid introducing
any movement artifacts in the scanner.
Consequently, one might argue that a motor
inhibition might also have led to activation of
the IFG region, particularly because both
factors (motor inhibition and mirror neuron
activity) may be associated with IFG activation
(Rowe and Siebner 2012; Bien et al. 2009).
However, we do not consider any possible
inhibition component to be more prominent
because the mirror component is essentially a
presumption for the other, and the overt
imitation of most mirror perceptions in healthy
adult humans is non-volitionally inhibited,
leading to covert imitation (Barkley 2001).
Nevertheless, it is impossible to differentiate
this definitely.
Another limitation may have been the
task-imminent inequality between our two sets of
dynamic stimuli, especially that concerning the
amount of biological motion. Whenever a task is
designed to provide differentiated stimuli in
this way, one cannot entirely exclude the
possibility that the extra activation in BA 9
under the yawning condition is merely due to
additional facial motions. Nevertheless, BA 9
has previously been reported to be active in
higher cognitive functioning (see above). The
small number of participants used here (11
total) might also be regarded as a limitation
because it did not allow us to perform
correlational analyses between the activation
and the contagions indicated by the
participants.
Via the MNS, physiological and associated
emotional states of two individuals can be
shared based on perceived motor patterns (Can et
al. 2003). This so-called motor empathy or
empathic resonance is one component within a
multi-component model of human empathy that is
adjacent to and underlies the development of
cognitive and emotional empathy (Gallese 2007;
Preston and de Waal 2002; Meltzoff and Decety
2003; Decety and Lamm 2006; Keysers and Gazzola
2007; Uddin et al. 2007; Blair 2005). Based on
our results, we conclude that a connection can
be demonstrated between the MNS and higher
cognitive empathic functions such as
mentalizing, as represented in the BA9.
In summary, we conclude that the easily
observable behavioral sign of CY is based on MNS
activity and, therefore, it can be considered an
expression of an individual's empathic
abilities. It would be interesting to study the
contagion effect besides the behavioral level,
utilizing functional imaging of patients with
impairments in their empathic abilities, such as
those with autism (Senju et al. 2007),
psychopathy (Hagenmuller et al. 2012), PTSD
(Nietlisbach et al. 2010), or schizophrenia
(Haker and Rössler 2009).