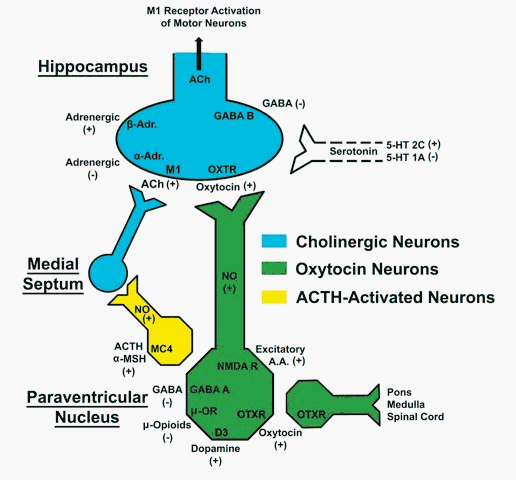
Oxytocin-induced
yawning: Sites of action in the brain and
interaction with mesolimbic/mesocortical and
incertohypothalamic dopaminergic neurons in male
rats
Sanna F, Argiolas A, Melis MR.
Department of Biomedical
Sciences, Neuroscience Section, and Centre of
Excellence for the Neurobiology of Addictions,
University of Cagliari, and Institute of
Neuroscience, National Research Council,
Cagliari, Italy.
Oxytocin (80ng) induces yawning when
injected into the caudal part of the ventral
tegmental area, the hippocampal ventral
subiculum and the posteromedial nucleus of the
amygdala of male rats.
The behavioural response occurred
concomitantly with an increase in the
concentration of extracellular dopamine and its
main metabolite 3,4-dihydroxyphenylacetic acid
(DOPAC) in the dialysate obtained from the shell
of the nucleus accumbens and of the prelimbic
medial prefrontal cortex by means of
intracerebral microdialysis. Both oxytocin
responses were significantly reduced by
d(CH(2))(5)Tyr(Me)(2)-Orn(8)-vasotocin, a
selective oxytocin receptor antagonist, injected
in the above brain areas 15min before oxytocin.
Similar results were obtained by activating
central oxytocinergic neurons originating in the
paraventricular nucleus of the hypothalamus and
projecting to the ventral tegmental area, the
hippocampus and the amygdala, with the dopamine
agonist apomorphine given at a dose that induces
yawning when injected into the paraventricular
nucleus.
Since oxytocin is considered a key regulator
of emotional and social reward that enhances
amygdala-dependent, socially reinforced learning
and emotional empathy, mesolimbic and
mesocortical dopamine neurons play a key role in
motivation and reward, and yawning in mammals is
considered a primitive, unconscious form of
empathy, the present results support the
hypothesis that oxytocinergic neurons
originating in the paraventricular nucleus of
the hypothalamus and projecting to the above
brain areas and mesolimbic and mesocortical
dopaminergic neurons participate in the complex
neural circuits that play a role in the above
mentioned functions.
Introduction
Yawning is a phylogenetically old,
stereotyped event that occurs alone or
associated with stretching and/or penile
erection with a low frequency in humans, in
animals from reptiles to birds and mammals,
under different conditions (see Argiolas and
Melis, 1998). Yawning occurs at bed and waking
times and in boring situations more frequently
than at other times of the day and situations.
In rats and non human primates yawning is
androgen-dependent and sexually dimorphic, with
males yawning more than females (see Argiolas
and Melis, 1998; Graves and Wallen, 2006).
The internal physiological stimuli that
evoke spontaneous yawning and its physiological
functions are unknown, although a role has been
suggested in stretching and arousal (Baenninger,
1997; Provine et al., 1987a), in normalizing
internal ear pressure (Laskiewicz, 1953), in
increasing oxygen-CO2 exchange in the lung
(Provine et al., 1987b), or even in cooling of
the brain (Gallup and Gallup, 2008) (but see
also Elo, 2010, 2011).
As yawning occurs concomitantly with an
increased cortical electroencephalographic
activity, it has been also suggested that
yawning is an ancestral vestige survived through
evolution that occurs when attention is low and
arousal needs to be increased (see Bertolini and
Gessa, 1981; Huston, 1971, but see also
Guggisberg et al., 2007). Yawning is also
contagious, as it can be also evoked in humans
or non-human primates by a yawn produced by
another subject of the same specie. This led to
suggest that yawning may be linked to empathy
(Anderson et al., 2004; Campbell and de Waal,
2011; Palagi et al., 2009; Paukner and Anderson,
2006).
Neuropharmacological studies revealed that
occurrence of yawning alone or associated with
stretching and penile erection, is under the
control of several neurotransmitters and
neuropeptides at central level. Of these, the
best known are adrenocorticotropin,
alpha-melanocyte stimulating hormone and related
peptides, acetylcholine, dopamine, serotonin,
excitatory amino acids, oxytocin,
gamma-amminobutyrric acid (GABA) and opioid
peptides. Some of these interact at the level of
the paraventricular nucleus of the hypothalamus
(PVN) in the control of this behavioural
response.
Briefly, dopamine receptor agonists,
N-methyl-D-aspartic acid (NMDA) and oxytocin
itself induce yawning by activating
oxytocinergic neurons projecting from the PVN to
extra-hypothalamic brain areas, such as the
hippocampus, the amygdala, the ventral tegmental
area (VTA), the frontal cortex, the nucleus
accumbens and/or the medulla oblongata, while
GABA and opioids prevent this behavioural
response induced by the above compounds by
inhibiting these neurons (see Argiolas and
Melis, 1998; Melis et al., 2007, 2009, 2010;
Succu et al., 2007).
Recent studies show that oxytocinergic
neurons reaching the VTA, the hippocampus and
the amygdala activate mesolimbic/mesocortical
dopaminergic neurons, and that the stimulation
of these dopaminergic neurons in turn activates
oxytocinergic neurons in the PVN, i.e., those
controlling erectile function and sexual
behaviour (see Melis et al., 2007, 2009; Succu
et al., 2007, 2011). This neural circuit
including oxytocin and mesolimbic/mesocortical
dopamine neurons, is thought to play a role in
the main aspects of both the anticipatory phase
(sexual motivation and reward) and the
consummatory phase (erection and copulation) of
sexual activity (see Melis and Argiolas,
2011).
As oxytocin is also considered a key
regulator of emotional and social behaviour in
both females and males (Lee et al., 2009;
Neumann, 2008 and references therein) or a
neuropeptide that determines the sexually
dimorphic aspect of social reciprocity and
enhances amygdala dependent, socially reinforced
learning and emotional empathy (Hurlemann et
al., 2010), and yawning may be an unconscious
signal that communicates one's own
physiological/ psychological state to other
members of a social group, e.g., a primitive,
unconscious form of empathy (Campbell and de
Waal, 2011; Guggisberg et al., 2010; Lehmann,
1979; Preston and de Waal, 2002), the above
neural circuit might also play a role in these
functions.
In order to test this possibility, we
studied: 1) the effect of oxytocin injected into
the VTA, the hippocampal ventral subiculum and
the posteromedial nucleus of the amygdala
(PMCo), on yawning and on the concentration of
extracellular dopamine in the shell of the
nucleus accumbens (NAs) and in the prelimbic
medial prefrontal cortex (mPFC) by means of
intracerebral microdialysis; and 2) if yawning
induced by the activation of PVN oxytocinergic
neurons projecting to the above brain areas by
the dopamine agonist apomorphine, occurs
concomitantly to an increase in extracellular
dopamine in the NAs and in the mPFC.
Discussion
The present results show that oxytocin (80
ng) induces yawning when injected into the
caudal VTA, the ventral subiculum of the
hippocampus or into the PMCo, three brain areas
containing oxytocin nerve endings (Adan et al.,
1995; Buijs, 1978) and rich in oxytocin
receptors (Boccia et al., 2007; Freund-Mercier
et al., 1987; Vaccari et al., 1998). This dose
of oxytocin is similar to those that induce
yawning and penile erection when the peptide is
injected into the CA1 field of the hippocampus
(Melis et al., 1986) or penile erection when the
peptide is injected into the caudal VTA (Melis
et al., 2007; Succu et al., 2008), the ventral
subiculum or the PMCo (Melis et al., 2009).
The pro-yawning effect of oxytocin did not
occur when the peptide was injected in
surrounding areas outside the VTA, the ventral
subiculum and/or the PMCo, strengthening the
anatomical specificity of the oxytocin effect
(see also Melis et al., 2007, 2009). As the
pro-yawning effect of oxytocin injected into the
VTA, the ventral subiculum or the PMCo was
markedly reduced by the prior injection of
d(CH2)5Tyr(Me)2-Orn8-vasotocin, a potent and
selective oxytocin receptor antagonist
(Bankowski et al., 1980), this effect is
apparently mediated by the stimulation of
oxytocin receptors present in these brain areas,
as found for oxytocin-induced yawning when
injected into the PVN (see Argiolas and Melis,
1998).
This study also shows that VTA, ventral
subiculum and PMCo oxytocin-induced yawning
occurs concomitantly to an increase in the
concentrations of extracellular dopamine and
DOPAC in the dialysates obtained from the NAs
and the mPFC. As expected, extracellular
dopamine and DOPAC increases were also markedly
reduced by the prior injection of a dose of
d(CH2)5Tyr(Me)2-Orn8-vasotocin that reduced
yawning, in line with the hypothesis that this
effect is also mediated by the stimulation of
oxytocin receptors present in these brain areas
(see above).
Dopamine and DOPAC increases in the NAs and
the mPFC dialysates and yawning were already
measurable within the first 15 min period after
oxytocin injection into the caudal VTA, but not
into the ventral subiculum or the PMCo, when
yawning occurred mainly in the second 15 min
period after oxytocin injection and the dopamine
increase.
This resembles the appearance of penile
erection, which was observed only 30 min after
injection of oxytocin into the ventral subiculum
or the PMCo (see Melis et al., 2007, 2009) and
is at variance from the effect of the peptide on
yawning and penile erection that appear in
5&endash;10 min after injection of the peptide
into the PVN (Melis et al., 1986). However,
since accurate assessment and comparison of the
temporal patterns of dopamine release and
yawning appearance is not feasible due the fact
that dialysate aliquots collection and yawning
episodes counting are made only at 15 min
intervals, it remains to be ascertained whether
this indicates that more steps than those
required for the dopamine increase need to be
activated after oxytocin injection in these
areas when compared to the VTA before yawning
may occur.
Despite this uncertainty, the increase of
extracellular dopamine and DOPAC in the
dialysate from the NAs and from the mPFC induced
by oxytocin injected into the VTA, the ventral
subiculum or into the PMCo and its reduction by
d(CH2)5Tyr(Me)2-Orn8-vasotocin, which blocks
oxytocin receptors, confirms that oxytocin
injected into these brain areas activates
mesolimbic and mesocortical dopaminergic
neurons.
This in turn leads to the appearance of
yawning by yet unknown mechanisms. The
activation of these dopaminergic neurons by
oxytocin injected into the VTA may occur
directly through oxytocinergic receptors located
in the cell bodies of dopaminergic neurons, as
already suggested for penile erection induced by
oxytocin injected into the VTA.
Accordingly, oxytocinergic fibres impinging
on VTA dopaminergic cell bodies retrogradely
labelled by Fluorogold previously injected into
the nucleus accumbens have been identified by
double labelling immunohistochemistry (Melis et
al., 2007). As to the mechanism by which
oxytocin injected into the ventral subiculum or
into the PMCo activates mesolimbic and
mesocortical dopaminergic neurons, this may
occur through the activation by the stimulation
of oxytocinergic receptors of direct or indirect
neuronal (mainly glutamatergic) pathways
projecting from the ventral subiculum or the
PMCo to the VTA, as already discussed in detail
(see Melis and Argiolas, 2011).
Indeed, oxytocin injected into the ventral
subiculum or the PMCo at a dose similar to that
used in this study induces not only yawning, but
also penile erection. This response was shown to
be secondary to an increase in NO production, at
least in the VTA and in the ventral subiculum
(Melis et al., 2010; Succu et al., 2008). While
NO in the VTA seems to activate directly
mesolimbic/ mesocortical dopaminergic neurons by
increasing cGMP production (Succu et al., 2008),
in the ventral subiculum NO stimulates glutamic
acid neurotransmission, leading to the
activation of neural efferent (possibly
glutamatergic) projections to yet unknown brain
areas, which increase glutamic acid
neurotransmission in the VTA, thereby modulating
the activity of mesolimbic and mesocortical
dopaminergic neurons (Melis et al., 2009, 2010;
Succu et al., 2011).
Irrespective of the exact mechanism through
which oxytocin injected into the VTA, the
ventral subiculum or the PMCo, causes the
activation of mesolimbic and mesocortical
dopaminergic neurons, this also leads to the
occurrence of yawning. One possibility is that
the activation of mesolimbic and/or mesocortical
dopaminergic neurons causes the activation of
incerto-hypothalamic dopaminergic neurons, which
impinge on PVN oxytocinergic neurons mediating
yawning, as already suggested for penile
erection induced by oxytocin injected into the
VTA, the hippocampal ventral subiculum or the
PMCo. Indeed oxytocin injected into the VTA
increases extracellular dopamine in the
dialysate obtained not only from the NAs, but
also from the PVN (Melis et al., 2007).
In line with this interpretation, this study
shows that a dose of apomorphine well known for
its ability to induce yawning when injected into
the PVN by stimulating dopamine D2-like
receptors located in the cell bodies of PVN
oxytocinergic neurons (see Sanna et al., 2011,
2012 and references therein), increases also
extracellular dopamine in the dialysate not only
from the NAs but also from the mPFC, and that
these responses are markedly reduced by
d(CH2)5Tyr(Me)2-Orn8-vasotocin given ICV (see
also Melis et al., 2007).
Although further experiments are necessary
to show that yawning occurs concomitantly to an
increase in extracellular dopamine not only in
the dialysate obtained from the NAs and the
mPFC, but also from the PVN, after the injection
of oxytocin into the ventral subiculum or into
the PMCo, together the above findings suggest
that oxytocinergic neurons originating in the
PVN and projecting to the VTA, the hippocampal
ventral subiculum and the PMCo, and mesolimbic
and mesocortical dopaminergic neurons
(originating in the VTA and projecting to the NA
and the mPFC) participate to a complex neural
circuit involved in the occurrence of yawning.
The involvement of the above oxytocinergic
and dopaminergic neurons in the occurrence of
yawning deserves some comment. Although the
internal physiological stimuli that evoke
spontaneous yawning and its physiological
functions are unknown (see Introduction), its
occurrence has been linked to the environment
(increased vigilance level, danger, hunting
prey) or even to communicative actions (sign of
aggressiveness, hierarchical dominance,
frustration, sexual excitement, boredom), or a
mean of synchronizing activities within the
group (see Giganti and Salzarulo, 2009;
Guggisberg et al., 2007, 2010; Senju, 2010).
Interestingly, brain oxytocin has been shown to
be important in coordinating well-defined
activities related to sociosexual behaviours and
to have anxiolytic and anti-stress properties
(see Baskerville and Douglas, 2010). Indeed
oxytocin receptor-null mice exhibit deficits in
social recognition and social memory, suggesting
a role of the neuropeptide in facilitating
interaction between individuals (Ferguson et
al., 2000, 2001). Similarly, disruption of
dopamine signalling in transgenic mice also
causes social abnormalities (Baskerville and
Douglas, 2010).
Common central brain regions involved in
mediating socio-sexual behaviours include the
nucleus accumbens, the prefrontal cortex, the
hippocampus, the amygdala, the VTA the medial
preoptic area (MPOA), the PVN and the supraoptic
nucleus (SON). All these limbic and hypothalamic
brain areas appear to be crucial components for
the execution of socio-affiliative behaviours in
rodents and for modulating neural reward
pathways as a consequence of social interaction
(Aragona et al., 2006; Edwards and Self, 2006;
Keverne and Curley, 2004; Young and Wang,
2004).
In particular, mesolimbic and mesocortical
dopaminergic neurons are thought to play a key
role in motivation and reward, and an increase
in extracellular dopamine concentration in the
dialysate from the NAs and from the mPFC has
been usually related to motivation and reward
processes (Everitt and Robbins, 2005; Wise and
Rompre, 1989) as well as to the execution of
goal-directed behaviours (Goto and Grace,
2005).
Therefore yawning might be considered as
a symptom that reflects the activation and/or
inhibition of brain areas important in the
control of social behaviour.
In this regard, it is noteworthy that a
lower number of yawning episodes (Lehmann, 1979)
and diminished plasma oxytocin levels have been
reported in schizophrenic patients with
neuroendocrine dysfunction and emotional
deficits (e.g., with predominant negative
symptoms) (Goldman et al., 2008) when compared
to healthy volunteers, even after apomorphine
administration (Legros et al., 1992). Likewise,
in male rats chronic treatment with
phencyclidine, an animal model of schizophrenia
(Javitt and Zukin, 1991), causes a decrease of
OT mRNA expression in the PVN and induces social
deficits which are reduced by oxytocin infused
into the amygdala (Lee et al., 2005), and
adjunctive intranasal oxytocin reduces symptoms
in schizophrenic patients (Feifel et al.,
2010).
Yawning is also altered in autism, where
contagious yawning is markedly reduced or absent
(Helt et al., 2010; Senju et al., 2007) as found
in schizophrenia (Haker and Rossler, 2009; see
Walusinski, 2009 for a review on yawning and
disease). The lack of contagious yawning seems
to occur with lower concentrations of plasma
oxytocin in autistic children when compared to
healthy controls (Modahl et al., 1998). Dopamine
dysfunctions have been also found in autism
(Gadow et al., 2008; but see also Anderson et
al., 2008), making it plausible a joint role for
oxytocin and dopamine in brain regions relevant
for this pathological condition, such as the
VTA, the nucleus accumbens, the hippocampus and
the prefrontal cortex (Baskerville and Douglas,
2010; Skuse and Gallagher, 2009).
In this regard, it is noteworthy that
functional magnetic resonance imaging (fMRI)
studies have shown that contagious yawning
activates the parahippocampal gyrus
(Schürmann et al., 2005), a part of the
limbic system including the amygdala and the
hippocampus, which are strictly connected to the
VTA, the nucleus accumbens and the mPFC, and the
ventromedial prefrontal cortex as well (Nahab et
al., 2009).
Finally, impaired social and emotional
capabilities are often observed in drug addicts
(Brady et al., 2007; van den Bosch and Verheul,
2007), and a huge body of experimental evidence
suggest that central dopamine and oxytocin
neural pathways may be two potential neural
correlates mediating drug addiction (Carson et
al., 2010; Everitt and Robbins, 2005; McGregor
et al., 2008).
In conclusion, oxytocin induces yawning not
only when injected into the PVN or the CA1 field
of the hippocampus, but also when injected into
the VTA, the hippocampal ventral subiculum and
the PMCo. This occurred with a concomitant
increase of extracellular dopamine and its main
metabolite DOPAC in the dialysate from the NAs
and the mPFC. Oxytocin-induced yawning and
extracellular dopamine increase in the NAs and
the mPFC are markedly reduced by
d(CH2)5Tyr(Me)2-Orn8-vasotocin, which blocks
central oxytocin receptors.
Similar results are obtained with
apomorphine injected into the PVN, a treatment
that activates oxytocinergic neurons projecting
to the VTA, the hippocampus and the amygdala
(Melis and Argiolas, 2011; Melis et al., 2007,
2009, 2010; Succu et al., 2007, 2008,
2011).
Also in this case yawning occurred
concomitantly with an increase of extracellular
dopamine and DOPAC in the NAS and mPFC
dialysate. These results confirm that PVN
oxytocinergic neurons projecting to the VTA, the
hippocampus and the amygdala activate directly
or indirectly mesolimbic and mesocortical
dopamine neurons, which in turn activate yet
unknown neural pathways leading to the
activation of incerto-hypothalamic dopamine
neurons impinging on the cell bodies of PVN
oxytocinergic neurons controlling yawning other
than erectile function and sexual behaviour
(Melis and Argiolas, 2011). This neural circuit
and the mechanisms described above may be also
involved in other functions in which dopamine
and oxytocin are thought to play a role, from
the regulation of emotional and social
behaviours (Baskerville and Douglas, 2010;
Everitt and Robbins, 2005; Lee et al.,
2009,2009; Neumann, 2008), to the control of
motivation and rewarding processes linked not
only to natural stimuli (feeding, sexual
behaviour) (Everitt and Robbins, 2005; Fulton,
2010) or to drugs of abuse (Carson et al., 2010;
Kovacs et al., 1998), but also to mental
pathologies, such as schizophrenia and autism
(see Baskerville and Douglas, 2010; Gadow et
al., 2008; Lee et al., 2009; McGregor et al.,
2008; Modahl et al., 1998).
Sanna F,
Succu S, Melis MR, Argiolas A. Dopamine
agonist-induced penile erection and yawning:
Differential role of D(2)-like receptor subtypes
and correlation with nitric oxide production in
the paraventricular nucleus of the hypothalamus
of male rats. Behav Brain Res.
2012;230:355-364