-
- -Giganti F, Hayes
MJ, Akilesh MR, Salzarulo P. Yawning and
behavioral states in premature infants. Dev
Psychobiol. 2002;41(3):289-96.
- -Giganti
F, Hayes MJ Cioni G, Salzarulo P Yawning
frequency and distribution in preterm and near
term infants assessed throughout 24-h recordings
Infant Behav & Development
2007;30(4):641-647
- -Giganti
F, Ziello ME Contagious and spontaneous
yawning in autistic and typically developing
children CPL 2009
- -Giganti
F, Zilli I. The daily time course of
contagious and spontaneous yawning among humans.
J Ethol 2011;29(2):215-216
- -Giganti
F, Zilli I, Aboudan S, Salzarulo P. Sleep,
sleepiness and yawning. Front Neurol Neurosci.
2010;28:42-6.
- -Giganti
F, Salzarulo P. Yawning throughout life.
Front Neurol Neurosci. 2010;28:26-31
- -Giganti
F, Toselli M, Ramat S. Developmental trends
in a social behaviour: contagious yawning in the
elderly. Giornale di Psicologia dello Sviluppo.
2012;101:111-117
- Giganti
F, Guidi S, Ramat S, Zilli I, Raglione LM, Sorbi
S, Salzarulo P. Yawning: A behavioural
marker of sleepiness in de novo PD patients.
Parkinsonism Relat Disord 2013
- -Zilli
I, Giganti F, Salzarulo P. Yawning in
morning and evening types. Physiol Behav
2007;91(2-3):218-222
- -Zilli
I, Giganti F, Uga V. Yawning and subjective
sleepiness in the ederly. J Sleep Res
2008;17:3003-308
- Tous
les articles consacrés au
bâillement
foetal
- Fetal
yawning: all
publications
-
-
- Human, adult observational studies have
established that yawning is a stereotyped action
pattern that occurs at low frequency throughout
the waking period (Provine,
1986a, 1986b). It is a complex behavior that is
characterized by gaping of the mouth, a long
inspiration followed by a shorter expiration.
Like other stereotyped patterns, when initiated,
it proceeds to completion with minimal influence
from sensory feedback. Yawning often is
accompanied by stretching, chewing, and
suppression of locomotor activity. The frequent
inclusion of whole body stretching and pende
erection in animal studies has led te, the
characterization of the behavior pattern as the
"stretching-yawning syndrome" (Argiolas &
Melis, 1998).
Yawning is facilitated by DA, acetylcholine,
serotonin, nitric oxide, and adrenocorticotropic
peptides which have been shown to stimulate
oxytocinergic, paraventricular nucleus neurons
and projections to hippocampus, pons, and
medulla. Central nervous system (CNS) injection
of adrenocorticotropic hormone (ACTH) or
alpha-melanocyte stimulating hormone and related
peptides increases yawning which continues for
several hours; induced yawning is suppressed by
opioid peptides and gamma aminobutyric acid
(Argiolas
& Melis, 1998; Ferrari, Gessa, &
Vargiu, 1963; Gessa, Pisano, Vargiu, Crabai,
& Ferrari, 1967).
In human adults, yawning bas been related
to a change in activity or arousal state
and is, therefore, tied to CNS arousal
modulation. Yawning increases in the hour before
bedtime and after waking (Provine,
Hamernik, & Curchack, 1987c) and when
subjects are watching or participating in
uneventful, repetitious activities (Provine
& Hamernik, 1986a). In a study of cab
drivers working long shifts, yawning increased
along with head, neck, and leg movements toward
the end of the shift in the early morning hours
(Sakai & Takahashi, 1975). Cortical
electroencephalographic activity (a marker of
increased arousal) increases during yawning
(Bertolini & Gessa, 1981). These findings
have been interpreted as evidence that yawning
may be potentiated when an increase in arousal
is needed to maintain alertness and vigilance in
a waking state.
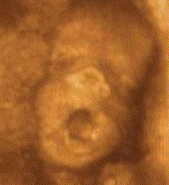
Yawning is a precocious behavior that is
observed sporadically in fetuses as early as 12
to 14 weeks of gestational age (De
Vries, Visser, & Prechtl, 1985). No
changes in the incidence of yawns between 20 and
36 weeks of gestational age have been observed
in the fetus by Roodenburg, Wladimiroff, van Es,
and Precht (1991). In full-term infants, yawns
were reported on the first day of life (Gesell,
1928). However, yawning in neonates is not
reported in studies of spontaneous behavior in
neonates, perhaps, in part, due to its
exceptionally low incidence (e.g., Korner, 1969,
1973; Korner & Beason, 1972). In the only
study to describe neonatal yawning in some
detail, Wolff (1987) observed that infant yawns
occur mainly near, or possibly anticipating, the
onset of sleep, together with reddening of the
eye sclera and drooping of the eyelids. State Il
sleep follows yawning with a mean latency of 7
min.
The present study examined yawning during
very early postnatal development in relation to
contextual behaviors and spontaneous alterations
in behavioral state. Since yawning is a
precocious behavior during early fetal life,
appearing many weeks before discemable
sleep-wake states, its expression gradually may
become linked to state. Further, since adult
yawning is increased during sleep-wake
transitions, we aimed to investigate if yawning
may be a corollary of state transition and/or
temporally close to sleep state during early
development. We also examined the temporal
association of yawning and the frequent bursts
of generalized activity expressed by the neonate
particularly during active sleep (AS) (Hayes,
Plante, Fielding, Kumar, &
Delivoria-Papadopoulos, 1994; Robertson, 1987).
[...]
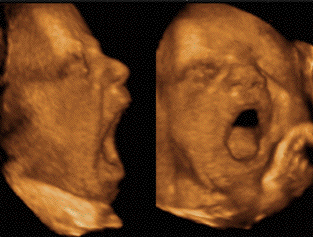 - RESULTS
Yawning Incidence : Spontaneous
yawning is rare. The rate of yawning across all
infants in the 5-hr nighttime recording period
averaged approximately one yawn/hr (median = 4.5
yawns/night). Table 3 shows that, using the
quotient statistic described earlier, there were
significant differences in the probabilities of
yawning across the behavioral states, Friedman's
X 2 (3) = 8. 0 1, p = .04. The highest
prevalence of yawning was in the D state. Using
the Wilcoxon test to compare all possible
pairings of behavioral states revealed that a
significantly lower quotient of yawning is
present in QS coinpared to W (p = .04), D (p =
.04), and AS (p = .04), No significant
differences were found for the other quotient
comparisons. Only 1 infant, the youngest (30
weeks of PCA), showed any yawning in QS.
Table 2. Coding Criteria
State Description
Active sleep (sommeil paradoxal) The
infant's eyes are closed with periodic REMs
occurring. Occasionally, eyes will be open
during REMs. Motor activity occurs in bursts of
10 to 60 s (generalized whole body movements
including facial, mouth, and head movements),
with muscle tone remaining low between movement
bures.Typically, respiration is irregular and
costal, which is evident by observing the thorax
and chest.
Quiet sleep(sommeil profond) The
infant's eyes are shut. Motor activity is low,
and muscle tone is moderate. Occasional mouthing
or sucking movements may occur. The infant's
respiration is even and abdominal in nature.
Wake (réveil) The infant's eyes
are open, focused (i.e., but not rapid REM
movements, often accompanied by head and arm
movements; whole sequence resembles an
orientation response), and scanning the
environment. State may include crying or
fussing. Motor activity may be high or low.
Respiration is regular and tracks motor
activity.
Drowse (endormissement) The infant's
eyes open and close slowly. When open, the eyes
are unfocused with a "heavy-lidded" appearance.
Characteristics of both sleep and wake states
are present, and respiration is even.
Yawn State : The distribution of sleep
states for the infants in this study was typical
for premature infants of this age (Giganti et
al., 2001-, Holditch-Davis & Thoman, 1990;
Meyers et al., 1998; AS: mean=48.9%, SD = 13.2;
QS: mean = 26.4l'/c, SD = 8.9; D: mean = 11. 1
%, SD = 6.4; W: mean = 12%, SD = 6.6). Nursing
interventions occurred on average for 4.8% (SD =
5.3) of the session.
To evaluate the temporal organization of
yawning and state, yawns in each state were
examined in relation to the preceding or
following state (Table 4). QS was not included
because there was only one example of a QS yawn
in the dataset. Each yawn state (e.g., AS, D, W
with at least one yawn) was examined for the
state of the preceding or following 3-min epoch.
The relative probability of the preceding or
following state was examined for its relation to
state stability or state change.
Preceding State : The preceding state
for each yawn state was compared To nonyawn
states from the same subjects. A two-way
cross-tabulation statistic was used to examine
the expected frequencies of each preceding state
given each yawn state. Preceding state
probabilities were found to be nonrandom and
predicted by the yawn state, X2 = 85.7, df = 12,
p <.000 1. Follow-up comparisons found that
yawn states AS, X2 = 16.2, df = 2, p<.0001,
or W, X = 11.6, df=2, p<.0001, were generally
preceded by the same state. That is, preceding
state and yawn state were stable. As shown in
Figure 1, the preceding state to D yawn state
was not stable, X2 = 2.6, df = 2, p <.27.
That is, for D yawn state, the preceding state
was either D (0.55) or W (0.40).
Next, AS, W, and D nonyawn states were
examined similarly. These data were drawn
randomly from the same sleep recording for each
subject as the yawn-state analysis. As was found
for the yawn states, nonyawn states predicted
preceding state, X2 = 98.2, df = 12, p <.000
1. AS and W nonyawns states were stable in the
preceding 3-min epoch, AS: X2 = 19.6, df=2,
p<.0001; W: 100% of all preceding states were
W. Unlike yawn D state, nonyawn D state was
stable, i.e., usually preceded by D state, X 2 =
6.25, df = 2, p <.04.
Following State : Following states for
yawn and nonyawn states were examined in the
same manner. Overall, yawn state predicted
following state, X2 = 108, df = 12, p <
.0001. Follow-up comparisons revealed that yawn
states predicted following states for all three
states, AS: X 2 = 28.9, df = 2, p <.00 1; W:
X2 = 7.36, df = 2, 2 p <.007; D: X = 6.4, df
= 2, p <.O 1. These findings indicate that,
unlike preceding states, the following states of
yawn D state are not characterized by state
change.
Nonyawn states also predicted following
states, X2 = 103, df= 12, p <.000 1.
Follow-up comparisons revealed that nonyawn
states predicted following states for AS, X 2 =
9.8, df = 1, p >.001, and W (100% of the
cases), but nonyawn D states were not stable, X2
= 3.25, df = 2, p <.20. As shown in Figure 2,
the probabilities for following states in
nonyawn D state are D=0.70, AS=0.20, and
W=0.10.
Contextual Behaviors : The type of
behaviors that were coded are defined in Table
5. These categories have been used previously to
categorize movement type in premature infants
under similar conditions (Hayes et al., 1994;
Hayes, Smith, Herrick, Roberts, & Swanson,
1997) and are inspired by previous work by
Smotherman and Robinson (1988).
In the analysis of contextual behaviors, the
30 s prior to and following each yawn were
examined and compared to nonyawn periods.
Nonyawn periods were matched to yawn periods for
subject and state type. The results show that
the 1 -min yawn and nonyawn periods matched for
state were different in the total frequency of
behaviors, yawn: X2 =17.33, df=4, p <.002;
nonyawn: X = 8.00, df = 4, p <.09. Yawn
periods were more behavior-rich than nonyawn
periods. This difference was in the total
frequency of behaviors only. It was not found
when examining before and after periods
separately or when the data were examined
according to state type.
The only finding specific for behavioral
typology was that facial movements were
proportionally more common in AS yawn versus
nonyawn 1-min contextual periods, X 2 = 7, df =
2, p <.03. D and W yawn states, as well as
nonyawn AS, W, or D state, showed no
différences in the proportion of separate
behavior types.
DISCUSSION
In premature infants, the rare behavior of
yawning is not randomly distributed with regard
to behavioral state across the night. Thirty to
35 PCA infants are Ilemergent" with regard to
state, Èe., rapidly developing the
capacity to express reliable and stable state
organization (Curzi-Dascalova, Peirano, &
MorelKahn, 1988; Parmalee, Wenner, Akiyama,
Schultz, & Stem, 1967). Nonetheless, yawns
were observed in all states except QS. This
result suggests that the necessary and
sufficient conditions for the elicitation of
yawns are not present, or may be actively
inhibited, during QS.
As has been observed in adults, yawning
prevalence in these infants showed a trend
towards highest levels during D state (Provine,
Tate, & Geldmacher, 1987b). One possibility
suggested by earlier work is that yawning is
seen during the D state and is associated with
efforts to remain awake. In animal studies,
yawning is associated with EEG activation and is
triggered by peptides such as ACTH. In this
study, our approach to examining the question of
the effect of yawning on ongoing states of
arousal was to examine yawning placement in the
strearn of state organization.
We found that preceding state stability was
temporally associated with yawn events in D
state, but not in AS or W states. When compared
to epochs of nonyawn D state, yawns in D state
had a higher incidence of unstable preceding
state. In yawn D state, the preceding state was
either D or W. This finding suggests that when
the preceding state is W state transitions to D
may be associated with an increased probability
of yawning.
Following states to a yawn state (whether D,
W, or AS) were characterized by state stability.
However, D state was unstable when yawns were
not present. Under these conditions, the
following state was either AS or W when state
change occurred. It is proposed that the absence
of yawning in D state may increase the
probability of a state transition in the
following 3-min epoch. The sequence analysis
suggests that some yawning effects are
state-specific. In D state, yawning is
associated with recent state change (i.e.,
"preceding" state instability), but state
stability in the following state epoch is sound
for all yawn states. Interestingly, al! nonyawn
states are stable for preceding state.
D state bas been viewed as a transitional
state between wake and sleep (Salzarulo, 1973;
Salzarulo, Giganti, Fagioli, & Ficca, in
press). Yawning may serve as an arousal
modulator in D that is potentiated by an
unstable preceding state. Yawning under these
conditions may increase arousal sufficiently to
stabilize the D state, which is consistent with
the position that nonyawn (but not yawn) D
epochs were unstable for the sequence of
following states. In total, these findings
suggest that D state is stabilized by yawn
events.
The temporal analysis of yawning revealed
that behavior rate was increased in the 1-min
window of yawns compared to nonyawn periods.
This result is reminiscent of the finding that
cab drivers yawned more and were more
behaviorally active at the end of their shift.
Both findings suggest that yawning is associated
with an increase in nonspecific motoric
activation.
Premature infants in this study were in AS
between 50 and 70% of the time in the 5-hr
recording period. W and D contextual behaviors
were similar in type to AS periods, although
lower in both frequency and variety. AS motor
bursts are characterized by a rich variety of
behaviors: stretching, facial movements, general
movements, and so on. The temporal window of
yawning in AS had more facial movements than AS
periods without a yawn. This result suggests a
spreading activation of facial motor patterning,
as has been suggested during movement analyses
in our earlier work (Hayes et al.,
1994).
-
-
-
 - Gianluca Ficca & Piero Salzarulo
"Lo Sbadiglio Dello Struzzo" ,
Bollati Boringhieri, Torino,
2002
|