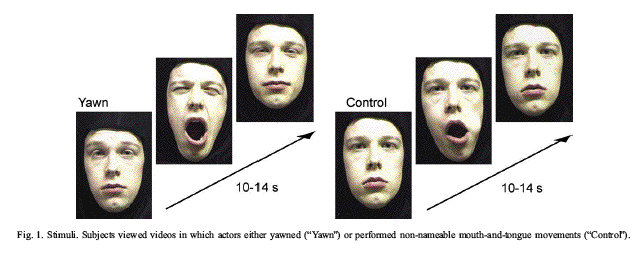
Yawning, an evolutionary old motor pattern
observed in many animals, is contagious, at
least in primates (Anderson
et al., 2004). Attempts to explain the
contagiousness of yawning as a result of, e.g.,
low oxygen or high carbon dioxide levels in a
shared environment, have proved unconvincing
(Provine, 1986;
Baenninger,
1997). Other explanations stress communicative
functions of yawns, e.g., by interpreting them
as social cues that synchronize group behaviour
(Deputte, 1994;
Daquin et al.,
2001). Such synchronization could be essential
for species survival and works without action
understanding, like when a flock of birds rises
to the air as soon as the first bird does so
supposably as it notices a predator.
We aimed at pinpointing the neural
correlates of yawn contagiousness by determining
those brain areas that are activated when
healthy adults observe other people yawn but do
not (yet) yawn themselves. We specifically
addressed the "mirror-neuron system"
(MNS) that is known to be activated while
subjects view another person's object-related
motor acts (Gallese
et al., 1996; Rizzolatti
et al., 1996, 2001; Hari et al., 1998; Decety
and Grezes, 1999; Nishitani and Hari, 2000,
2002; Buccino et al., 2001; Iacoboni et al.,
2001; Rizzolatti and Craighero, 2004).
This system, especially the posteroinferior
frontal cortex (Broca's area in the left
hemisphere), is considered to specifically
support action perception and understanding as a
prerequisite for "true imitation", i.e., the
copying of goal-directed actions when an
individual learns some part of a new behavior
(Rizzolatti et al., 2001; Wohlschlger et al.,
2003). The action-related MNS, and a
corresponding visceromotor mirroring system for
shared sensory and emotional experience, are
thought to provide the neuronal framework for
insight into other minds, even to the level of
empathy (Gallese et al., 2004).
We hypothesized that MNS is less likely to
be activated by stereotypical motor patterns
that are not "truly" imitated but are rather
triggered automatically. We thus expected
observation of yawns to activate brain areas
involved in the perception and evaluation of
orofacial gestures without additional activation
in the inferior frontal cortex because of the
high stereotypy of the individual yawning
patterns. To identify the neural correlates of
yawn contagion, we acquired functional MR images
from 30 adult volunteers who watched videotaped
sequences of yawns and control stimuli. The
subjects were instructed to attentively view the
faces but yawning was not mentioned in any way.
The subjects were prevented from overt yawning
by constraining their heads and chins to avoid
movement artifacts. The effectiveness of these
constraints was confirmed by post-experimental
questionnaire in which none of the subject reprt
overt yawning.
[...]
Results and discussion
Yawn videos evoked a stronger tendency to
yawn than did control videos (subjects' ratings
on the 1Ð5 scale, mean F SEM 2.8 F 0.2 vs.
1.4 F 0.1, respectively; P b 0.001, Wilcoxon).
The difference between both ratings, i.e., our
measure of individual yawn susceptibility, was
insignificantly higher for female than for male
subjects (1.5 F 0.5 vs. 1.2 F 0.3; n.s., P =
0.57, t test). Subjects also reported a stronger
tendency to imitate yawns than mouth movements
in control videos (2.1 F 0.3 vs. 1.4 F 0.2; P b
0.005, Wilcoxon). Contrasting the blood oxygen
level dependent (BOLD) signals for observing
yawn videos vs. baseline (blank screen), we
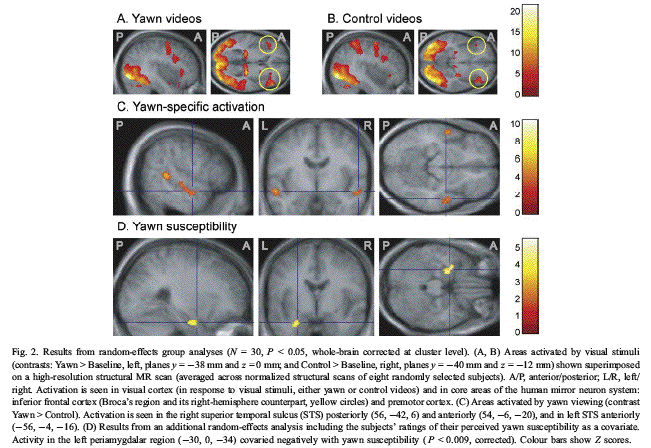
found a pattern of activation that included
inferior frontal cortex and premotor cortex
(Fig. 2A, P b 0.05, corrected at the cluster
level, random-effects group analysis involving
all voxels of the brain).
A similar pattern of activation was found
for control videos vs. baseline. The observed
reactivity to both types of facial stimuli in
the inferior frontal cortex (Broca's
région and its tight hemisphere
counterpart) and in the premotor cortex, i.e, in
the core areas of the human MNS, agrees with
earlier work (Decety and Greezes, 1999;
Rizzolatti et al., 2001; Nishitani and Hari,
2002; Buccino et al., 2004; Rizzolatti and
Craighero, 2004).
Contrasting yawn vs. control videos,
significant activations were found in the medial
visual cortex (Yawn > Control) and in the
lateral visual cortex (Control > Yawn). One
possible explanation for this difference is that
actors' eyes were closed during part of the yawn
videos but not during the control videos.
Moreover, parietal and premotor activations in
the Control > Yawn contrast could indicate
that subjects followed the actors' complex and
unpredictable tongue movements. Robust Yawn >
Control differences were found in the posterior
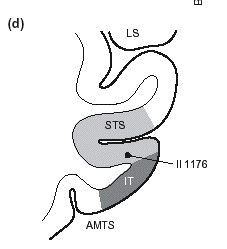
part of the right superior temporal sulcus (STS;
local cluster maximum in MNI coordinates at x =
56, y = 42, z = 6; local Zmax = 4.98; see Fig.
2C) and in the anterior parts of STS bilaterally
(x = 56, y = 4, z = 16; Zmax = 4.70 and x = 54,
y = 6, z = 20; Zmax = 4.02).
This activation of the posterior part of STS
agrees with the established selectivity of the
STS for processing socially relevant cues in the
perception of biological motion in general, and
of faces in particular (Perrett and Mistlin,
1990; Allison et al., 2000). STS also has a role
in the detection of the goals and outcomes of an
agent's behaviour (Frith and Frith, 1999;
Gallagher and Frith, 2003). In this context, it
could be argued that the tongue movements in the
control videos were more goal-directed than were
the yawns.
However, this difference could not explain
our pattern of STS activation, which was more
intense during yawns than during control videos.
Activation of the anterior part of STS was
maximal within 1 cm of a location where
intracranial event-related potentials have
indicated specificity for facial movements
compared with static faces (Puce and Allison,
1999). Although the human STS region is not
activated by self-paced execution of motor acts,
a necessary condition for an area to be
considered a part of the motor MNS, STS is an
important node during the typical activation
sequence seen during observation and imitation
of orofacial gestures (Nishitani and Hari,
2002).
In contrast, no suprathreshold activation
was detected in the Yawn > Control contrast
either in the inferior frontal cortex (Broca's
region or its right-hemisphere counterpart) or
in primary motor cortex; however, these areas
were clearly activated by both Yawns and Control
stimuli in our experiment. Despite the high
sensitivity in our study on N = 30 subjects,
random-effects analysis did not show Yawn vs.
Control differences in these regions. Moreover,
with 95% confidence, the Bayesian analysis
(Friston and Penny, 2003) ruled out that the
difference in activation between Yawn and
Control in Broca's region or primary motor
cortex would exceed a negligible 0.1% of the
global mean BOLD signal. The lack of
yawn-specific activation of Broca's region
supports our hypothesis: As the yawn
contagion relies on the release of a highly
stereotypical motor pattern rather than on true
imitation, yawn observation activates only a
subset of the brain areas that support action
understanding as a prerequisite for
imitation. Even during contagious yawns, the
details of another person's yawn are not
imitated.
Importantly, the above analysis indicated
that STS activation was evoked by the
observation of Yawns vc. control stimuli as
such, regardless of the participants'
subjectively perceived need to yawn. To identify
brain regions where the strength of this
differential activation would vary with
subjective yawn susceptibility, a second
random-effects analysis on the Yawn > Control
contrast images included the subjects' ratings
of yawning tendency as a covariate. A
statistically significant negative covariance
was observed between the subjects' ratings and
the Yawn-Control difference in the BOLD signal
from the left periamygdalar region (local
maximum at 30, 0, 34; Zmax = 4.68; P = 0.009,
corrected; Fig. 2D).
No regions of statistically significant
negative covariance were found in the
corresponding analysis for Control-Yawn
differences. In an additional random-effects
analysis, we verified that the covariance of
amygdalar activation with yawning tendency was
specific to the YawnÐControl contrast:
Although both YawnÐ Baseline or
ControlÐBaseline contrasts showed strong
amygdalar activation per se, the amygdalar
effect did not covary in these contrasts with
the subjects' ratings.
Furthermore, the periamygdalar site of
covariance did not show up in the
YawnÐControl contrast; thus, the covariance
result cannot be explained by differences
between yawn and control stimuli as such.
Periamygdalar activation has been associated
with the emotional load of social cues,
particularly those related to human faces
(Critchley et al., 2000; Phelps et al., 2000;
Tillfors et al., 2001; Winston et al., 2002). We
accordingly suggest that the observed negative
covariance between yawn susceptibility and
periamygdalar activation might reflect a
relationship between the effectiveness of yawn
contagion and implicit evaluation of facial
expressions. Such processing is known to occur
during the perception of faces even when it is
not relevant to the task (Critchley et al.,
2000; Phelps et al., 2000) or to accompany the
assessment of trustworthiness (Winston et al.,
2002). An alternative interpretation is based on
individual differences in social perception and
attribution of mental states; in a recent
behavioural study (Platek et al., 2003), such
differences correlated with the susceptibility
to yawn by contagion. However, these
explanations remain speculative before more
empirical data are available, and we cannot yet
provide a causal explanation for the observed
relation between amygdalar activation during
yawn viewing and the subjective tendency to
yawn. Nevertheless, this finding represents the
first known neurophysiological signature of
perceived yawn contagiousness.
In summary, our study on the neural
correlates of attentive viewing of other
persons' yawns results in three main
conclusions: (i) STS activation appears to
differentiate viewing of stereotypical yawns
from viewing of physically similar non-yawn
orofacial gestures, (ii) the absence of
activation in Broca's region and its
right-hemisphere homologue, important parts of
the MNS, in the YawnÐControl comparison
speaks for the non-imitative nature of the yawn
contagion that can occur without detailed action
understanding, and (iii) the negative covariance
between the subjective yawn susceptibility and
the differential amygdalar activity (meaning
that perceived contagiousness increases as
amygdalar activation decreases) suggests a
relationship between the effectiveness of yawn
contagion and the face-processingrelated
emotional analysis during social
interaction.
References
1. Perrett et al. (1992) Phil. Trans. B 355:
23-30.
2. Bonda et al. (1996) J. Neurosci. 16:
3737-3744.