- The ethogram
-
- Research began with a qualitative
description of the behaviour shown by members of
the field and laboratory colonies. It soon
became apparent that the various activities
carried out by these fish were but a series of
numerous, rather discrete, species-specific,
behavioural units which were sufficiently
stereotyped in form, duration and orientation,
that their occurrence could easily be noted by
trained observers. These units, in turn,
provided the basic material from which arose all
subsequent analyses, interpretations and
experimental verification of certain points of
the study.
-
- There were three basic types of be units:
motor patterns (i.e. locomotory movements and
postures), colour patterns; and sounds. Although
colour patterns varied in duration, motor
patterns and sounds were considered
instantaneous events since the duration of a
given event was generally less than I s (there
were exceptions, e.g. Chasing, Skimming,
HeadStanding; but even these were usually only a
few seconds in duration). All patterns,
irrespective of type, entered the ethogram since
all were important in reaching an understanding
of the intricate mechanisms that controlled the
behaviour of the subject species.
-
- Behaviour patterns within the ethogram were
not analysed, quantitatively, as to their
respective variations in intensity or duration
(as carried out, for example, by Rasa 1969, on
the Lateral display of Pomacentrus jenkinsi).
Though certainly possible, such analyses would
have expended much time and effort for a minimum
of return since patterns, at the level of our
analysis, by and large, demonstrated Typical
Intensity (Morris 1957; see discussion by Barlow
1963). I agree with the interpretations by
Baerends (1957) and Nelson (l964a) that the
elementary action patterns described in the
ethogram, though complex, were relatively
stereotyped and it appeared self-evident and
natural to describe and record behaviour in
terms of activities of that order. I do not
claim that these action patterns (motor, sonic
and colour) correspond to Fixed Action Patterns
(Lorenz 1956) since descriptions were based
largely on convenience and not upon the criteria
of the F.A.P.
-
- Any ethogram necessitates some type of
ordering. This was done, mainly, by grouping
action patterns together under various titles,
each title having an apparent functional
correlate (e.g. courtship, agonism, nest care,
etc.). I realize fully the danger of such
categorization since often morphological and
contextual criteria result in hybrid
classification (see discussion by Nelson 19
64b). Nevertheless, impressions gained during
the qualitative phase formed a basis for
classification and a test of the reality and
significance of this classification was an
important part of the subsequent quantitative
phase of the study. Fortunately, the bicolor
damselfish exhibited many discrete patterns. 1f
it had shown but a few consistent patterns, such
a classification would have been meaningless,
except on the basis of given probabilities that
a single event occurs within several or all
functional categories.
-
- Motor Patterns Patterns and
Postures of Basic Locomotion
-
- Fish of the field and laboratory colonies
demonstrated the same basic locomotory movements
and postures. Movements were often so rapid,
however, that adequate descriptions were often
accomplished only by reduced-speed
cinematography and video-taping.
-
- Forward Swimming was accomplished largely by
the caudal fin with the pectoral fins being used
mainly for changing direction. This mode of
swimming, though common to the genus,
Eupoinacentrus (Emery 1968) is contrasted with
the 'pectoral swimming' of two other well-known
genera of damselfishes, i.e. Abudefduf: Cummings
1968; Helfrich 1958; Stevenson 1963; and
Chromis: Abel 1961; Myrberg, Brahy & Emery
1967; Turner & Ebert 1962. The caudal fin is
generally used by the latter genera only during
sudden and rapid manoeuvring. Fins were expanded
to varying degrees during slow Forward Swimming,
the degree of expansion depending on the
swimming speed and the particular motor pattern
being exhibited.
-
- Backward Swimming was uncommon in bicolors,
but a number of specific situations predictably
brought it to the fore: (1) a territorial holder
approaching an unfamiliar, small object at, or
near, its territory (e.g. a new conspecific, a
congeneric, a hand, a rod); (2) a female
approaching closely a territorial holder outside
its territory; and finally (3) rearward
movements during Head-Standing and Fluttering.
Backward Swimming was accomplished by the
pectoral fins paddling alternately while the
caudal fin and the posterior portion of the
dorsal fin were used as rudders.
-
- Hovering occurred as the fish remained
motionless in the water column. The pectoral
fins performed the major sculling movements
while the rear portion of the dorsal and tip of
the caudal fin provided apparent compensation.
The median and pelvic fins were slightly
expanded. The pattern was uncommon since the
bicolor was an active fish; however, it occurred
most frequently following Nipping activity.
Certain motor patterns appeared similar to
Hovering but maximal spreading of median fins
and the degree of spreading by the pelvic fins
(maximal-Fin-Spread; minimal-Lateraland
Frontal-Display) readily distinguished them as
being separate and distinct.
-
- Fin-Spread (Fig. 6) was a basic locomotory
posture often evident during an agonistic
encounter or while carrying out braking
movements, following a rapid manoeuvre. The
median and pelvic fins were maximally spread (as
were the pectorals during braking movements).
The body was horizontal and occasionally, the
rear portion of the body had a slight lateral
bending. This posture is the same as noted in
other fishes (e.g. anabantids: Miller & Hall
1968; badids: Barlow 1962a; centrarchids:
Carter-Miller 1963; cichlids: Baerends &
Baerends-van Roon 1950; cyprinodonts: Foster
1967). A detailed account of this motor pattern,
applicable to the present species is found in
Myrberg (1965).
-
- Head-Stand was another basic locomotory
posture that was often evident during
interspecific interaction (e.g. when large
fishes such as snappers, grunts, groupers and
parrotfishes approached the nest of a guardian
male). The body was inclined, head downward; all
fins were spread maximally; the actor might be
standing still or moving slightly forward or
backward. The angle of inclination varied from a
slight pitch to over 60° from the
horizontal, the angle depending on the distance
between the object of attention and the actor.
If an intruder turned away, the actor, then,
rapidly regained a horizontal attitude and
carried on other activities. If the intruder,
however, moved within an apparent critical
distance, the actor suddenly 'exploded' from the
Head-Stand posture, lunging at its adversary
with great speed. Such explosive swimming and
the ram or bite that followed usually resulted
in the large adversary shaking itself and then
moving slowly away. Head-Stand was not often
followed by explosive swimming but a large
proportion of the occurrences ol explosive
swimming were preceded by Head Stand. Head-Stand
was rarely seen durinl intraspecific
interactions; rather its general re striction to
interspecific agonism placed it rathei clearly
apart from the numerous agonistk patterns to be
described later. The hydrodynamk aspects of this
posture, applicable to the present species, are
discussed by Myrberg (1965).
-
- Wandering was slow or moderate swimminl
about its environs by a fish that was neither
interacting directly with other fishes nor
showinl a recognizable motor pattern. This
activity, though non-descript, provided, if
nothing else, a slight pause in a hectic
recording session, the latter including as many
as 600 distinct patterns recorded from a single
individual during a hr, observation period. The
duration of Wandering though not considered in
this report, was obtained during recording
sessions since its significance to certain
problems has been amply demonstrated (see Barlow
1964).
-
 - Maintenance Patterns
-
- Included here are those motor patterns which
appeared to aid fish by either removing foreign
matter from the body (Chafing, Fin-Flicking,
Defecating, Body-Shaking) or providing a unique
type of muscular activity (Yawning, Tail-Down,
Tail-Dig). These patterns appeared to occur
independently of a direct social context,
feeding or nesting. Yet, they certainly did not
appear to form a single grouping based on a
common causal factor(s). This was borne out
later during the quantitative phase of the
study.
-
- Chafe was carried out against some hard
object. A fish swam to a plastic tube
(laboratory) or a frond of Sargassum (field) and
just prior to reaching the object, it
accelerated slightly. The fish then rubbed part
of its body (usually the mid-lateral section)
against the object with its fins fully expanded.
The subsequent rebound from the object was
either straight ahead or arching, the latter
resulting in circling back to the object. Two to
four Chafes often occurred together. Bicolors
rarely Chafed against the sandy bottom or
scoured rock, apparently preferring objects of a
smoother texture.
-
- Fin-Flick was often associated with Chafing.
While swimming about, a fish occasionally folded
and then expanded rapidly one or more of its
fins, primarily the dorsal and pelvics. The
different combinations of fins involved were
difficult to observe and since the pattern was
uncommon, we recorded simply the occurrence of
the general pattern rather than attempting to
dissect it into its possible variations.
-
- Yawn
involved a rapid opening of the mouth, often
maximally, accompanied by an equally rapid
expansion and subsequent lowering of the median
and pelvic fins. Although the pattern was easily
recorded from members of the laboratory colony,
it was difficult to record in the field. It
could be easily seen when using the full
telephoto capability of the UTY camera, but
recording sessions rarely employed that aspect
of the camera, since it restricted the area of
surveillance.
-
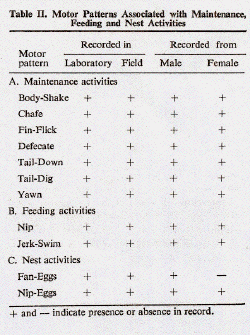 -
- Tail-Down. A fish swimming near the
substrate would suddenly stop, within a few
millimetres of the substrate (also folding its
fins at the same time), and then drop its caudal
fin to the substrate. Following this manoeuvre,
the fish would either regain a horizontal
position and swim off or follow through with the
next pattern.
-
- Tail-Dig involved a rapid beating of the
caudal fin at the substrate while the actor
moved forward, body inclined about 20°
(headup) for a distance of about 10 cm. During
this short traverse, the anal, pelvics and
spinus dorsal were folded against the body; the
pectorals, though held close to the body,
appeared to act as stabilizers, as did the rear
portion of the dorsal fin. The sweeping action
of the caudal fin resulted in substantial sand
being blown from the area of traverse and it
appeared initially to be a technique whereby
food matter was cast into the water column. Yet
we never recorded a bicolor turning around after
completing Tail-Digging and then feeding or even
'inspecting his work'. Males dug in their
territories and along their borders, but it was
also recorded occasionally some distance away
from such areas. The function of depressions
left by such activity remained a mystery. They
were never used for any clear purpose. I have
observed Tail-Digging in numerous species of
damselfishes and its wide-spread occurrence
indicates an important functional basis. I wish
to venture a suggestion that this pattern has
some, as yet unknown, chemical function related
to nest activity or territoriality in the
various species that perform it. Centrarchids
(Lepomis) also show a pattern quite similar to
Tail-Dig (Carter-Miller 1963), and there it
appears as a nest directed activity.
-
- Body-Shake was rarely seen in bicolors. The
pattern consisted of a lateral shaking of the
entire body, two or three times in rapid
succession. The fins were folded slightly. The
pattern occurred so seldom during recording
sessions that subsequent analysis of it was not
possible. It did not appear, however, to have
any function other than possibly removing
foreign matter from the body.
-
- A comparative summary of the apparent
maintenance patterns of bicolors showed that all
patterns were carried out by members of the
laboratory and field colonies, irrespective of
sex.
-
 -
|

