Stimulation,
control and phylogenetic projection of the
teleostean yawn reflex
FH McCutcheon
Department of Animal
Biology, School of Veterinary Medicine,
University of Pennsylvania,
Philadelphia
Introduction
Psychological features of sighs and yawns in
man, including the yawn's "infectious" character
(Ganong, 1965), have long been remarked in
poetic and psychiatric literature. The function
of these impositions on the rhythmic ventilation
process remains obscure. The physiological role
of sighs has attracted some recent attention
with the study of pulmonary surfactants
(Clements, 1962) and emphasis on normal alveolar
distension, especially in small mammals
(McCutcheon, 1953; Agostoni et al., 1959).
The role of yawns has been described in
animal behavior as psychological "displacement
activity", after yawns became a noteworthy
element in the studies of courtship and brooding
behavior of birds and especially fishes
(Forselius, 1957). The physiological role of
yawns appears to have attracted attention,
however, only in a few species of fishes, the
sea bass (Centropristes striatus) and the
pinfish (Lagodon rhomboides) in particular
(McCutcheon, 1958, 1966). The yawn reflex has
received little attention in any terrestrial
animal although yawns do occur in every class of
vertebrates. In addition to those represented by
the literature already cited, namely Mammalia,
Ayes and Teleostij, my personal, observations
include yawns in Reptilia (i.e. blacksnake,
Coluber constrictor, and chameleon, Anolis
carolinensis) and Amphibia (i.e. bullfrog
tadpole, Rana catesbiana). This reflex may be
absent in modern adult amphibians. More than
5000 frogs, toads and salamanders, including
about fifteen species, have been used in our
laboratories for various physiological
experiments among which some yawns should have
been noted or recorded if, in fact, the reflex
existed in these animals. Not a single instance
of yawns has been observed.
The overt features of the yawn reflex in
fishes are like those in man, dog or cat; there
is wide mouth gaping, body tensing and often
isometric stretching or extension of appendages.
Yawns can be triggered in fishes by direct
manipulation of the swimbladder volume through a
syringe and needle, or indirectly by
manipulation of the ambient pressure
(McCutcheon, 1958, 1966). The specific receptors
and stimulus characteristics are not known.
Yawns are correlated with the processes of
swimbladder volume adjustment; that is, with the
secretion-absorption and, most immediately,
compression of gas. These processes can respond
to less than 10 cm H20 pressure in the pinfish
(McCutcheon, 1966). They compensate for volume
changes which are persistent with reference to a
control or "fix" level of buoyancy. However,
yawns also occur in exceptional species of
fishes which lack a swimbladder. Some local sea
mullet or southern whiting (Menticirrhus
americanus) recently added to our laboratory
stock have provided our first example, and
Magnuson & Prescott (1966) reported yawns in
the Pacific bonito (Sarda chiliensis). Yawns in
such fishes may involve the buoyancy effect of
gases normally present in the intestine
(Scholander, 1954), but no data are available on
this subject.
Confirmation is here reported for earlier
observations about the stimulating effect of
imposed negative pressure on the yawn reflex in
pinfish (McCutcheon, 1966). Also, evidence is
provided from electrical stimulation of the
pinfish brain for an extra-medullary "yawn
center".
Materials and methods
Ambient pressure stimulation
The Philadelphia Aquarama provided the means
to study two species of physocistous marine
fishes which are typically very large (18,000
vs. 100 g) and unavailable in-shore when
compared to the pinfish and sea bass previously
reported. One display tank at the Aquarama,
holding about 21,000 l of sea water, contained
five jewfish (Promicrops itaiara), which were
74-76 cm standard length and weighed an
estimated 16-18 kg, and two red snappers
(Lutianus aya), which were 59-61 cm in length
and weighed 11-13 kg. Two gray snappers
(Lutianus griseus) and a schoolmaster (Lutianus
apodus) completed the display of teleosts in the
tank; these were enough like the red snappers to
require no separate mention. All of these fishes
were habituated to the aquarium where they had
been maintained in the same tank more than 2
years. The control depth of water in the tank
was 122 cm. The normal water circulation and
filtering system allowed this depth to be
readily altered and restored for our test
purposes. It was reduced 12 cm in 5 min when a
stimulus for the yawn reflex was required.
Electrical stimulation
A stanchion-like restraint and holder were
made to secure the head of a pinfish in a fixed
position. This provided for stereotaxic
placement of a stimulating electrode the brain
for a survey using conventional
neurophysiological technique. The fish's brain
was exposed under O 2% urethane anesthesia. The
fish breathed normally in slightly urethanized
aerated sea water during brain preparation and
preliminary stimulation. Aeration was stpped
when depressed respiration was required. A
stainless wire, 0,02 cm in diameter served as a
unipolar stimulating electrode. An electronic
research stimulator provided current in a
circuit calibrated and monitored with a
cathode-ray oscilloscope.current in a circuit
calibrated and monitored with a cathode-ray
oscilloscope.
Twenty pinfish, 14,1-15,5 cm standard
length, were examined. Stimuli of 0,5 msec
duration, 20-60/sec frequency and 0-3-1 5-0 V
were used. Various sites were stimulated,
following a grid pattern, at successive 0,1 mm
depths in each brain. Histological methods for
post-mortem identification of the electrode path
were not used in these exploratory studies. As
the stimulation sites were tested moving
posteriorly from the forebrain, unitary
responses of various effectors, including upper
jaw, lower jaw, operculum, eye and fin muscles,
were elicited with 0,9-1,5 V, until the
cerebellum was reached. Here a very low
threshold, massive, lateral flick or thrusting
movement of the body and tail was induced by as
little as 0,3 V single shocks, a threshold
one-third or less of the previous responses.
Eventually, in some experiments of long duration
in which respiration failed, the flick became
depressed. Its threshold increased so that
stimuli of O,5-2V no longer elicited a flick,
but other responses which had been precluded by
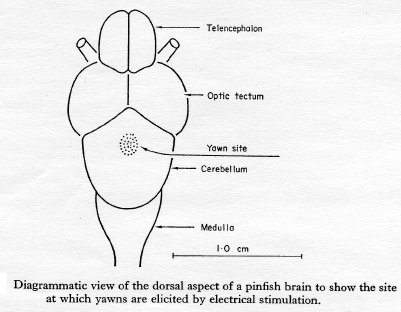
the flick began to appear. A site was found
under these conditions in which massive yawns
were induced by a stimulus of 1,6 V.This region
(Fig. 1) was then explored in five fish, during
a period without aeration of the sea water.
Ambient pressure stimulation
The control behavior and pressure-sensitive
reactions of the jewfish and snappers were like
those of the sea bass and pinfish. They each had
a specific territorial site. This included the
same "fix0 position" feature identified in
pinfish studies; that is, a discrete place
occupied with remarkable precision (less than 2
min variation in pinfish) when critical buoyancy
test-adjust activity was in progress. In this
vicinity the yawns occurred. The jewflsh, which
is bottom oriented like the sea bass, remained
resting on the bottom near the fix0 posit
sometimes for an hour or more. The snapper,
which like the pinfish cruises free away from
the bottom, held its fix0 position only briefly
and occasionally a pause from slow swimming
about the tank. During periods of immobilization
the fix0 position, whenever a yawn was about to
occur, the dorsal fin flicked rapidly and then
stood fully erect momentarily, thus signalling
the end of a test-adj episode in the same way as
in the sea bass and pinfish. In these larger
fishes a notably in the jewfish, which has an
unusually conspicuous mouth, the yawn its was
even more prolonged and its consummatory
character more evident.
It now seems proper to consider the yawn
reflex a basic functional characteric of
teleosts in general. The phylogenetic
persistence of this reflex through amp bians and
reptiles to birds and mammals, taken together
with the homology of the swimbladder and lung,
make the central control of yawns of special
comparative interest. Consequently the brain of
the pinfish was examined for a possible "yawn
center".
Electrical stimulation
Within the region indicated on Fig. 1, full
mouth gaping, abducted opercule and body-wall
contraction were induced by tetanic stimulation
with stimuli 0,5 msec duration at 60/sec
frequency. The lowest effective strength in this
circonstans was 0,85 V at 6,0 mm depth. The
dorso-ventral depth of the cerebellum was ab 8,0
mm at this site. Current of 15 V would elicit a
yawn at 20 mm depth. However, dorsal fin
erection, a part of the normal yawn complex, did
not occur until 4-6 V were applied at 3-6 mm
depth.
A general muscular contraction appeared in
the thoraco-lumbar body region during yawns.
Progressive increase in wall tension, with
marked displacement the scales over the
swimbladder region, was evident during
increasing stimulati of the yawn site. To
confirm this movement as actual compression, a
needle was inserted in the swimbladder of one
preparation and connected to a small wall
manometer. Stimulation by 10 V increments gave
successive pressures of 0,8; 3,5; 4,4; 7,2 and
l2 mm H20.
Swimbladder pressure variations are normal
concomitants of buoyancy change and adjustments
in fishes. Some studies have been reported on
the central control of buoyancy which are
consistent with a pressure-related cerebellar
yawn since Long (1959) explored the brain of the
carp (Cyprinus carplo) to determine if the
pressure control site could be identified. He
found a region of about 10 mm2 the caudal part
of the tecta optica where he could, by
electrical stimulation, elicited a 10-15 mm Hg
pressure increase in the swimbladder. He
demonstrated that compression was effected by
body wall rather than swimbladder musculat Yawns
were not a subject of investigation at that
time. Bianki (1964) investI the role of the
cerebellum in buoyancy by combining brain
extirpation with conditioning experiments in the
carp. He found that the cerebellum, but not
forebrain, was necessary for the development of
a stable conditioned reflex pressure changes in
the swimbladder.
MeduIlary respiratory centers, basic to the
breathing cycles, are well known in fishes
HeaIey, 1957) and in every other vertebrate
class. But extra-medullary respiratory centers,
pneumotaxic and apneustic, are well known only
in mammals, their normal role remains obscure
(Lambertsen, 1961). The extra-medullary,
apneustic-like character of the yawn site in
pinfish suggests the possibility that an
analogus role exists for the "apneustic center"
in mammals; namely, to function as a "yawn
center". As such, stimulated by a persistent
variation from some control level of thoracic or
pulmonary proprioceptive discharge, the center
would act to divert the ventilation mechanism
into a yawn sequence. This would serve restore
proprioceptive tension, reset the control level
and concomitantly stabilize the pulmonary
residual air volume. The latter is important
especially in those terrestrial animals with an
alveolar lung in an open system, namely birds
and mammaIs. Such an adaptation of the
teleostean yawn reflex for a modified
homeostatic role in terrestrial animals would be
consistent with the way in which the
phylogenetic development of breathing itself
(McCutcheon, 1964) proceeded among vertebrates.
Its confirmation will depend on elucidation of
the yawn reflex especiaIly in reptiles and
mammals.