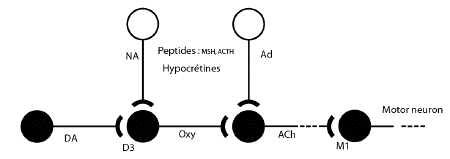
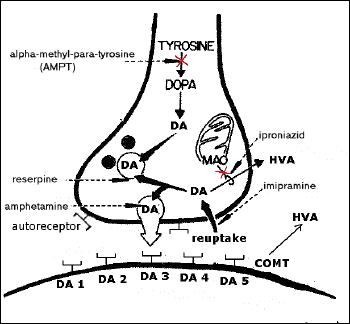
Dopamine
Agonist-Induced Yawning in Rats: A Dopamine D3
Receptor Mediated Behavior
Collins GT, Witkin JM, Newman AH, Svensson
KA, Grundt P, Cao J, Woods JH.
University of Michigan
Medical School
Abstract :
A specific role for the dopamine D3 receptor
in behavior has yet to be elucidated. We now
report that dopamine D2/D3 agonists elicit
dose-dependent yawning behavior in rats,
resulting in an inverted U-shaped dose-response
curve. A series of experiments was directed
toward the hypothesis that the induction of
yawning is a D3 receptor mediated effect, while
the inhibition of the yawning observed at higher
doses is due to competing D2 receptor
activity.
We compared several dopaminergic agonists
with a range of in vitro D3 selectivity,
including; PD-128,907, PD-128,908, quinelorane,
pramipexole, 7-OH-DPAT, quinpirole,
bromocriptine, and apomorphine with respect to
their ability to induce yawning in rats. A
series of D2/D3 antagonists differing in
selectivity for D3 over D2 receptors were
evaluated for their ability to alter the effects
of the dopamine agonists.
The antagonists L-741,626, haloperidol,
nafadotride, U99194, SB-277011A, and PG01037
were used to determine effects on dose-response
curves for D2/D3 agonist-induced yawning. In
addition, the potential contribution of
cholinergic and/or serotonergic mechanisms to
the yawning response was investigated using a
series of pharmacological tools including
scopolamine, mianserin, and the D3-preferring
antagonists; nafadotride, U99194, SB-277011A,
and PG01037 to differentially modulate yawning
induced by the PD-128,907, physostigmine, and
TFMPP.
The results of these experiments provide
convergent evidence that dopamine D2/D3
agonist-induced yawning is a D3 agonist mediated
behavior, with subsequent inhibition of yawning
being driven by competing D2 agonist activity.
Thus, dopamine agonist-induced yawning may
represent an in vivo method for selectively
identifying D3 and D2 receptor-mediated
activities.
Introduction:
Dopamine D3 receptors have received
considerable interest since originally cloned.
The D3 receptor shares significant sequence
homology with the dopamine D2 receptor, but
displays a much more restricted, limbic pattern
of distribution compared to that of the D2
receptor in the rat and human brain. Based in
large part on this restricted distribution and
high sequence homology, it has been hypothesized
that the D3 receptor may be of interest as a
pharmacological target for antipsychotics and
antiparkinsonian therapeutics. Additionally, the
D3 receptor is thought to play a role in
reinforcement pathways, as the D3 receptor is
expressed in high levels within the mesolimbic
dopaminergic system, and more specifically, the
nucleus accumbens shell.
However, progress in defining a role for the
D3 receptor has been slowed by the inability to
identify behavioral effects that can be linked
exclusively to a D3 mechanism. This is, at least
in part, due to the lack of pharmacologically
selective compounds acting at either the D3 or
D2 receptors, as well as the fact that
potentially selective agonists have failed to
elicit obvious, direct behavioral changes. While
D2/D3 agonists and antagonists have been shown
to produce changes in body temperature,
locomotor activity, and other behavioral
measures, a role for the D3 receptor in the
regulation of these effects has typically not
been confirmed by studies using D3 receptor
deficient mice. Recently, increases in locomotor
activity by MK801 and blockade of the convulsant
effects of dopamine uptake inhibitors have been
proposed as in vivo models of D3 receptor
activation. However, systematic replication of
those findings or confirmation by other means
has not been reported. The studies reported
herein provide evidence supporting the
contention that yawning induced by D2/D3
agonists is mediated specifically through D3
receptor activation.
The ability of dopaminergic agonists to
elicit biphasic yawning resulting in an inverted
Ushaped dose-response curve in rats has been a
long-studied phenomenon. An early hypothesis
regarding the biphasic regulation of
apomorphine-induced yawning behavior attributed
the induction of yawning behavior to a D2
agonist activity, while the inhibition seen at
higher doses was thought to be due to a
competing D1 agonist activation. The cloning of
the dopamine D3 receptor and the development of
agonists such as PD-128,907 and 7-OH-DPAT as
well as antagonists including U99194, SB-27701A
and PGD1D37 with greater degrees of in vitro
selectivity for the D3 receptor have allowed
greater insights into the regulation of
dopaminergic agonist-induced yawning behavior to
be made. Based on a series of studies examining
the unconditioned behavioral effects of
7-OH-DPAT, as well as binding studies
hypothesized in an extensive review that, D2/D3
agonist-induced yawning may be a D3 agonist
mediated effect, while the inhibition seen at
higher doses was a result of concomitant D2
agonist activation.
This hypothesis was evaluated in the present
studies using a host of pharmacological tools.
The abilities of a series of compounds with
varying in vitro selectivities for the D3
relative to D2 receptors to elicit yawning were
examined. A series of antagonists, again defined
by binding in vitro selectivity for the D3 and
D2 receptors, were evaluated with respect to
their modification of dose-response
relationships for D2/D3 agonists, with the
majority of the studies using PD-128,907 as a
prototype D3 agonist.
Finally, in addition to dopaminergic
mechanisms, yawning can be induced by
cholinergic or serotonergic compounds. While the
exact mechanisms and neural pathways involved in
the regulation of yawning behavior have not been
fully elucidated, there is a large set of data
that suggests that dopaminergic, serotonergic,
and cholinergic induction of yawning occur via
distinct mechanisms. In addition both
dopaminergic and serotonergic pathways are
thought to eventually feed onto cholinergic
neurons, thus allowing for differential
regulation of dopaminergic and serotonergic
yawning, with a cholinergic component common in
all three pathways. Therefore, some D3
antagonists that reduced PD-128,907 induced
yawning were also assessed for their capacity to
alter non-dopaminergicinduced yawning.
The convergent evidence from the agonist and
antagonist studies support the hypothesis that
dopamine agonist-induced yawning is mediated
specifically through activation of D3 receptors.
Therefore, yawning in rats may provide a
critical model for establishing the in vivo
activities of putative D3 selective ligands, a
first step toward defining their role in normal
and pathological physiological states.
[...]
Discussion:
Evidence has been provided in the present
paper to support the hypothesis that D2/D3
agonist-induced yawning behavior in rats is
mediated by agonist activation of the dopamine
D3 receptor, while the inhibition of yawning is
a result of a competing agonist activation of
the dopamine D2 receptor. ln agreement with the
majority of previous studies, all of the D2/D3
agonists tested with exception of bromocriptine
and PD-128,908, the inactive enantiomer of
PD-128,907, elicited significant, dose-dependent
increases in yawning behavior with inhibition
seen at higher doses, resulting in the
characteristic inverted U-shaped dose response
curve for yawning in rats. Evidence is also
provided for the selective antagonism of the
induction of yawning behavior by D3 preferring
antagonists, and the inhibition of yawning by
D2-preferring antagonists. In addition, the
current studies demonstrate that inhibition of
D3 agonist-induced yawning by D3-preferring
antagonists is a result of their selective
antagonist activity at the D3 receptor, and not
through antagonist effects at D2, serotonergic,
or muscarinic cholinergic receptors.
Yawning is a D3 mediated
behavior.
Several lines of evidence have been provided
in support of the hypothesis that yawning is a
D3 agonist mediated behavior. In general, all
D3-preferring D2/D3 agonists induced significant
amounts of yawning at low doses. While there
were no significant differences in the
effectiveness of the agonists with respect to
induction of yawning behavior with the exception
of apomorphine, there were differences in the
potency of the D2/D3 agonists to induce yawning.
The rank-order potency of the D2/D3 agonists to
elicit yawning behavior was as follows;
quinelorane, apomorphine, quinpirole, 7-OH-DPAT,
pramipexole, and PD-128,907, while bromocriptine
and PD-128,908 failed to elicit significant
levels of yawning. The stereoselectivity of the
yawning response with regard to PD-128,907
[and PD-128,908] is an important
finding, as dopamine receptors are selective
with respect to more rigid agonists. Taken
together with the findings of Stahle and
Ungerstedt (1984), who showed that (+)-3-PPP,
but not (-)-3PPP, will elicit yawning, our
current findings provide further evidence that
D2/D3 agonists are inducing yawning via
dopaminergic agonist mechanisms. Differences in
yawning induced by bromocriptine may be a result
of pharmacokinetic differences, as bromocriptine
has been shown to induce significant levels of
yawning in studies using a 60 minute observation
period.
Antagonists with a high degree of
selectivity for the D3 compared to the D2
receptor selectively antagonized the induction
of yawning behavior. Three of the four D3
preferring antagonists (U99194, SB-277011A, and
PG01037) tested in the current studies possess
the ability to dose-dependently and selectively
antagonize the induction of yawning by
PD-128,907, while having no effect on the
inhibition of yawning observed at higher doses.
As shown in figures 3G and 3D respectively,
SB-27701 IA and PG01037, D3-preferring
antagonists with similarly high degrees of in
vitro D3 selectivity (100- and 133-fold
respectively) produced almost identical effects
on PD-128,907 induced yawning; significant,
dose-dependent, downward/rightward shifts of the
ascending limb of the yawning dose-response
curve were observed, while the descending limb
of the dose-response curve for
PD-128,907-induced yawning was not changed.
Similar effects were seen with the moderately
selective (30-fold) D3-preferring antagonist
U99194, however, unlike SB-27701 lA and PG01037,
at relatively high-dose of 10.0 mg/kg, U99194
completely inhibited PD-128,907-induced yawning;
however, it should be noted that at this dose
U99194 effectively antagonized not only
dopaminergic, but cholinergic and serotonergic
yawning as well. Nafadotride, the least
selective (3-fold) of the D3-preferring
antagonists, was the only D3 antagonist to
produce a non-selective antagonism of yawning
behavior; shifting both the ascending and
descending limbs of the dose-response curve for
PD-128,907-induced yawning at the highest dose
tested. This effect was similar to that observed
with haloperidol, a non-selective dopamine
antagonist, and suggests that at a dose of 0.32
mg/kg, nafadotride is no longer selective for
the D3 receptor, but rather is active at both
the D3 and D2 receptors. Taken together, these
data provide strong support for the hypothesis
that the induction of yawning by D21D3 agonists
is mediated by an agonist activation of the D3
receptor.
Inhibition of yawning is
a D2 mediated effect.
We have also provided evidence in support of
the hypothesis that inhibition of D2/D3
agonist-induced yawning occuring at higher doses
is mediated by an agonist activity at the D2
receptor. As shown in figure 2A and 2B, the
D2-preferring antagonist L-741,626, at the first
behaviorally active dose (1.0 mg/kg),
selectively antagonized the inhibitory effects
of high doses of PD-128,907 and quinelorane,
resulting in a rightward shift in the descending
limbs while having virtually no effect on the
ascending limbs of the doseresponse curves for
both PD-128,907- and quinelorane-induced
yawning. In addition L741,626 produced a
rightward shift in the maximal effective dose of
P0-128,907 and quinelorane, resulting in an
increased effectiveness for both agonists. These
data not only suggest that L-741,626, at a dose
of 1.0 mg/kg, is an effective D2 antagonist in
vivo, but that it is also devoid of D3
antagonist activity.
Further support for the differential
regulation of yawning behavior by the D3 and D2
receptors was provided by the effects of the
non-selective DA antagonist haloperidol. As D3-
and D2-preferring antagonists selectively
antagonize the ascending and descending limbs of
the dose-response curve for D2/D3
agonist-induced yawning respectively, it would
be expected that antagonists with mixed D2/D3
actions, such as haloperidod, would shift both
the ascending and descending limbs of yawning
dose-response curves at their initial active
doses. Indeed, at the first behaviorally active
dose (0.032 mg/kg), haloperidol produced
rightward shifts in both the ascending and
descending limbs of the dose-response curves for
both PD-128,907- and quinelorane-induced
yawning. This not only suggests that the effects
of D3- and D2-prefemng antagonists are a result
of selective antagonist activity, but that
non-selective D2/D3 antagonists produce effects
distinct from those of other dopaminergic
antagonists on D3 agonist induced yawning.
However, it should be noted that in addition
to possessing high affinities for the D3 and D2
receptors, halopendol also has significant
affinities for the D1, D4, and D5 receptors. It
is, however, unlikely that activity at these
receptors is influencing PD-128,907-induced
yawning behavior as the D1/D5-selective
antagonist, SCH 23390, and the D4-selective
antagonist, L-745,870, at behaviorally active
doses did not alter yawning elicited by either
low (0.032-0.1 mg/kg) or high (0.32 mg/kg) doses
of PD-128,907. This provides further evidence
that D2/D3 agonist-induced yawning behavior is
under the direct control of the D3 (induction)
and D2 (inhibition) receptors, but not the D1,
D4, or D5 receptors. However, the possibility
remains that other dopaminergic receptors may
modulate D3 agonist-induced yawning elicited by
other D2/D3 agonists, as several of the agonists
tested, such as apomorphine, quinelorane, and
quinpirole, possess significant affinities for
the Dl, D4, and D5 receptors (apomorphine), or
04 receptor (quinelorane and quinpirole) in
addition to the D3 and D2 receptors.
Dopaminergic,
serotonergic, and cholinergic regulation of
yawning.
The findings of the current study confirm,
and extend those of earlier studies, and
demonstrate that while scopolamine will
dosedependently antagonize yawning induced by
cholinergic, serotonergic, and dopaminergic
agonists, serotonergic and dopaminergic
antagonists are able to selectively antagonize
yawning elicited by their respective agonists.
More specifically, nafadotride, SB-27701 lA, and
PG01037, D3-preferring antagonists with a wide
range (3-133 fold) of selectivities for the D3
receptor over the D2 receptor in vitro, were
able to selectively antagonize
PD-128,907-induced yawning, while having no
effect on yawning elicited by either
physostigmine or TFMPP . This suggests that
SB-27701 lA and PG01037 are not only selective
for the D3 over the D2 receptor, but that they
are also selective for the D3 receptor over
certain serotonergic and cholinergic receptors
at doses up to 56.0 mg/kg. Similarly, while
nafadotnde demonstrated little or no preference
for the D3 compared to the D2 receptor in vivo,
no serotonergic or tholinergic antagonist
activity was detected at doses up to 1.0 mg/kg.
However, in contrast to the effects of the other
D3-preferring antagonists, U99194, at a dose of
10.0 mg/kg, significantly antagonized yawning
elicited by PD-128,907, TFMPP and physostigmine,
suggesting that at higher doses, it is no longer
selective for dopaminergic receptors. While
U99194 is unique in this regard within this
group of D3-preferring antagonists, clozapine,
an antagonist with significant affinities for
dopaminergic, serotonergic and cholinergic
receptors has also been shown to antagonize both
dopaminergic and cholinergic yawning, suggesting
that antagonism of physostigmine-induced yawning
may be a reliable measure of anti-cholinergic
activity. Further evidence of an in vivo
anti-muscarinic activity of U99194 has been
demonstrated by Gaudie and colleagues (2001) who
showed in discrimination studies that U99194
generalized to a scopolamine cue, suggesting
that U99194 may possess anti-cholinergic
activity at higher doses. Although it has been
suggested that U99194 functions as a D3
selective antagonist in vivo at doses ranging
from 13.0 to 400 mg/kg based on its inability to
increase plasma prolactin, to induce catalepsy,
and to inhibit the induction of hypothermia by
PD-128,907, the results of the current study
suggest that while U99194 may be selective for
the D3 compared to the D2, a significant
anti-cholinergic effect is apparent at 10.0
mg/kg. Thus the current studies support the
hypothesis that dopaminergic, serotonergic and
cholinergic agonists induce yawning via distinct
mechanisms, and furthermore that yawning induced
by D2/D3 agonists is a result of agonist
activation of D3 receptors, and not serotonergic
or cholinergic receptors.
To summarize the results of the studies
reported herein, evidence has been provided in
support of the hypothesis that the induction of
yawning by D2/D3 agonists is mediated through an
agonist activity at the D3 receptor, while the
subsequent inhibition of yawning seen at higher
doses is a result of an increasing D2 agonist
activity. Based on these findings several
conclusions can be drawn: First, the ascending
limb of the doseresponse curves corresponds to
doses that are selectively activating D3
receptors over D2 receptors, while the
descending limb corresponds to those activating
both the D3 and D2 receptors. Additionally,
determinations of in vivo D3 potency and
effectiveness may be possible, based on the
onset and maximal amount of yawning elicited.
Furthermore, inhibition of yawning may provide
useful information regarding in vivo D2 potency,
and lastly, the shape of the dose-response
curves may allow for determinations of in vivo
D3 selectivity of D3-preferring D2/D3 agonists
to be made. The results of the current set of
studies have demonstrated that D3 selective
antagonism will only shift the ascending limb of
the yawning dose-response curve, that D2
selective antagonism will only shift the
descending limb of the yawning dose-response
curve, while non-selective D2/D3 antagonism will
shift both the ascending and descending limbs of
the dose-response curve for D2/D3
agonist-induced yawning behavior in rats. In
conclusion, as the current studies have provided
evidence that the induction of yawning behavior
by D2/D3 agonists is mediated by the D3
receptor, yawning may be an important
pharmacological effect that can be used in the
characterization, classification, and discovery
of in vivo D3 agonist and antagonist actions.
Thus, it may be possible to relate other
behavioral effects of D2/D3 agonists and
antagonists to their ability to modulate
yawning. Whether the potency and selectivity
measures of these compounds can be utilized
across behavioral measures will need to be
explored in the future.
-Collins
C, Truccone A et al. Pro-erectile Effects of
Dopamine D2-like Agonists are Mediated by the D3
Receptor in Rats and Mice. J J Pharmacol Exp
Ther 2009
-Canales JJ,
Iversen SD Psychomotor activating effects
mediated by dopamine D2 and D3 receptors in the
nucleus accumbens Pharmacol Biochem Behav 2000;
67;161-168
-Canales
JJ, Iversen SD Dynamic dopamine receptor
interactions in the core and shell of nucleus
accumbens differentially coordinate the
expression of unconditioned motor behaviors
Synapse 2000; 36; 297-306
-Collins GT, JM
Witkin et al Dopamine agonist-induced
yawning in rats: a dopamine d3 receptor mediated
behavior. J Pharmacol Exp Ther
2005;314(1):310-319
-Collins GT, Newman
AH,Woods JH et al.Yawning and hypothermia in
rats: effects of dopamine D3 and D2 agonists and
antagonists. Psychopharmacology
(Berl).
2007;193(2):159-170
-Collins GT. et
al. Food restriction alters
pramipexole-induced yawning, hypothermia, and
locomotor activity in rats: Evidence for
sensitization of dopamine D2 receptor-mediated
effects. JEPT 2008;325:691-697
-Collins
GT et al. Narrowing in on compulsions:
dopamine receptor functions Exp Clin
Psychopharmacol 2008,16(4):498,502
-Chen J, Gregory T.
Collins, et al. Design, Synthesis, and
Evaluation of Potent and Selective Ligands for
the Dopamine 3 (D3) Receptor with a Novel in
Vivo Behavioral Profile. J Med. Chem.,
2008;51(19):5905&endash;5908
-Collins GT,
Truong YN, et al. Behavioral sensitization
to cocaine in rats: evidence for temporal
differences in dopamine D(3) and D (2) receptor
sensitivity. Psychopharmacology (Berl).
2011;215(4):609-20.
Li SM,
Collins GT et al. Yawning and locomotor
behavior induced by dopamine receptor agonists
in mice and rat. Behav Pharmacol. 2010; 21(3):
171&endash;181.
-Banks RJ,
Mozley L, Dourish CT. The angiotensin
converting enzyme inhibitors captopril and
enalapril inhibit apomorphine-induced oral
stereotypy in the rat. Neuroscience.
1994;58(4):799-805
-Stoessl
AJ, Dourish CT, Iversen SD
Apomorphine-induced yawning in rats is abolished
by bilateral 6-hydroxydopamine lesions of the
substantia nigra Psychopharmacol; 1987; 93;
336-342
-Stoessl
AJ Effects of ageing on the behavioural
responses to dopamine agonists: decreased
yawning and locomotion, but increased stereotypy
Brain Research 1989; 495; 20-30
-Stoessl AJ,
Dourish CT, Iversen SD The NK-3 tachykinin
agonist senktide elicits yawning and chewing
mouth movements following subcutaneous
administration in the rat. Evidence for
cholinergic mediation. Psychopharmacology
(Berl). 1988; 95; 4; 502-506
-Cooper SJ;
Dourish CT Neural basis of drug induced
yawning in neurobiology of stereotyped behaviour
Oxford ed 1990
-Cooper SJ,
Rusk IN, Barber DJ. Yawning Induced by the
Selective Dopamine D2 Agonist N-0437 is Blocked
by the Selective Doparnine Autoreceptor
Antagonist (+)-UH 232. Physiol Behav.
1989;45:1263-1266
-Dourish
CT, SJ Cooper Yawning elicited by sytemic
and intrastriatal injection of piribedil and
apomorphine in the rat Pyschopharmacology 1985;
86; 175-181
-Dourish CT,
PH Hutson Bilateral lesions of the striatum
induced with 6-hydroxydopamine abolish
apomorphine-induced yawning in rats
Neuropharmacology 1985; 24; 11; 1051-1055
Based on studies in the
rat, Sokoloff et al. have made the valuable
suggestion that the D3 receptor is a
particularly important target for antipsychotics
in the mesolimbic DA system. These study in the
human demonstrates that the distribution of D3
receptors and D3 mRNA-bearing neurons is
consistent with relative segregation of the D3
subtype to the limbic striatum as well as it
primary and secondary targets and many sources
of its afferents.