The selective NK-3 tachykinin receptor
agonist senktide elicited yawning, chewing mouth
movements and sexual arousal following
subcutaneous administration (0.1-1.0 mg/kg) in
the rat. These responses were not significantly
affected by the dopamine antagonist haloperidol
(0.03 mg/kg) or by 6-hydroxydopamine lesions of
the nigrostriatal projection. In contrast, the
behaviours were markedly attenuated by the
peripheral and central muscarinic antagonist
scopolamine (1 mg/kg), but not by the peripheral
muscarinic antagonist N-methylscopolamine (1
mg/kg). These findings suggest that stimulation
of NK-3 receptors produces yawning, chewing and
sexual arousal by directly activating central
cholinergic neurons.
Introduction
The precise mechanisms underlying the
yawning response remain undetermined. Yawning
and stretching are elicited by a variety of
peptide hormones, including adrenocorticotrophic
hormone (ACTH), alpha-melanocyte stimulating
hormone (alpha MSH), oxytocin and prolactin
(Ferrari 1958; Gessa et al. 1967; Laping and
Ramirez 1986; Meus et al. 1986). Cholinergic
agents also induce yawning as well as vacuous
chewing mouth movements (Urba-Holmgren et al.
1977; Yamada and Furukawa 1980; Ushijima et al.
1984; Salamone et al. 1986). Finally, low doses
of dopamine agofists injected peripherally
(Yamada and Furukawa 1980; Mogilnicka and Klimek
1977; Urba-Holmgren et al. 1982; Protais et al.
1983; Gower et al. 1984; Mogilnicka et al. 1984;
Stahle and Ungerstedt 1984), intrastriatally
(Dourish et al. 1985) or into the
paraventricular nucleus of the hypothalamus
(Melis et al. 1987) also result in yawning and
sexual arousal.
It has been suggested that dopamine
agonist-induced yawning reflects the
disinhibiton of striatal cholinergic
interneurons secondary to autoreceptor-mediated
inhibition of nigrostriatal dopamine neurons
(Mogilnicka and Klimek 1977; Yamada and Furukawa
1980; Urba-Holmgren et al. 1982; Protais et al.
1983; Gower et al. 1984; Mogilnicka et al. 1984;
Stahle and Ungerstedt
1984; Dourish et al. 1985). While arguments
against the autoreceptor hypothesis have been
forwarded (Morelli et al. 1986; Serra et al.
1986; Longoni et al. 1987; Melis et al. 1987;
Serra et al. 1987), the evidence supporting this
hypothesis is at least two-fold: (I) The dose
response curve for dopamine agonist-induced
yawning is bell-shaped, with maximal response
rates occurring at doses which are selective for
autoreceptor stimulation (Yamada and Furukawa
1980; Urba-Holmgren et al. 1982; Protais et al.
1983; Mogiinicka et al. 1984; Stahle and
Ungerstedt 1984); (ii) Dopamine agonist-induced
yawning is markedly attenuated by bilateral
6-hydroxydopamine (6-OHDA) lesions of the
striatum (Dourish and Hutson 1985) or the
substantia nigra (Stoessl et al. 1987 a; Laping
and Ramirez 1988).
Senktide is a synthetic peptide which is
highly selective for the NK-3 tachykinin
receptor subtype (Wormser et al. 1986). We have
previously demonstrated that intracisternal or
subcutaneous injection of senktide induces
5-hydroxytryptamine (5-HT)-mediated behaviour
(head twitches or wet dog shakes, forepaw
treading, flattened body posture, Straub tail)
in mice and rats (Stoessl et al. 1987b; 1988).
We now demonstrate that yawning, chewing mouth
movements and sexual arousal are also induced by
peripheral administration of low doses of
senktide. These behaviours appear to be
cholinergically mediated, but independent of
dopamine autoreceptors, and provide evidence for
a tachykinin-cholinergic link in the central
nervous system.
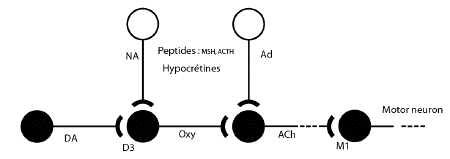
Discussion
These data indicate that activation of NK-3
tachykinin receptors elicits yawning, chewing
mouth movements and sexual arousal in rats. The
centrally active muscarinic antagonist
scopolamine, but not the peripheral muscarinic
antagonist N-methylscopolamine, blocked the
induction by senktide of all of these
behaviours. This suggests that these responses
are mediated by activation of central
cholinergic neurons. The cholinergic neurons
involved may be located in the striatum, in view
of the apparent striatal mediation of dopamine
agonist-induced yawning (Dourish et al. 1985)
and the known dopaminergic-cholinergic link in
drug-induced yawning (Yamada and Furukawa 1980).
However, the density of NK-3 binding sites in
the striatum is quite low (Beaujouan et al.
1986; Buck et al. 1986; Danks et al. 1986).
Another possible site of action is the
paraventricular nucleus of the hypothalamus.
This region is also thought to be important for
the expression of yawning (Melis et al. 1986;
Argiolas et al. 1987; Melis et al. 1987) and
displays dense NK-3 binding (Beaujouan et al.
1986; Buck et al. 1986; Danks et al. 1986), but
the presence of cholinergic neurons here is
controversial (see Kasa 1986 for review).
In vitro pharmacological studies indicate
that senktideinduced ileat contraction is
dependent on muscarinic transmission (Wormser et
al. 1986). Taken together with our observations,
this suggests that cholinergic neurons in both
the peripheral and central nervous system may
have NK-3 receptors which can act to stimulate
the release of acetylcholine.
The yawning induced by low doses of dopamine
agonists is also blocked by scopolamine (Yamada
and Furukawa 1980; Ushijima et al. 1984; Dourish
et al. 1985). However, the failure of
haloperidol (at a dose which has been shown to
block dopamine agonist-induced yawning; see
Gower et al. 1984; Mogilnicka et al. 1984;
Ushijima et al. 1984; Dourish et al. 1985) or
6-OHDA nigrostriatal lesions to significantly
attenuate senktide-induced yawning or chewing
mouth movements suggests that these effects of
senktide may occur independently of dopamine
autoreceptor stimulation. Although it may be
argued that the degree of striatal dopamine
depletion resulting from our lesions (64%) was
insufficient to allow this conclusion to be
drawn, a similar degree of depletion was
sufficient to block apomorphine-induced yawning
(Stoessl et al. 1987a). More importantly, there
was no relationship between striatal dopamine
levels in lesioned animals and the frequency of
senktide-induced yawning, so that animals with
marked depletions had high yawning rates, while
other animals with normal dopamine levels had
very low response rates.
It is noteworthy that the dose-response
curve for senktide-induced yawning is
bell-shaped. Two possibilities might explain the
disappearance of yawning at higher doses of
senktide. The first is behavioural response
competition at higher doses, a constellation of
other behavioural responses (i.e. wet dog
shakes, forepaw treading, etc.) interfere with
yawning. The second is pharmacological
antagonism, in that serotonergic stimulation
(which appears to result from senktide
administration) attenuates the yawning induced
by low doses of dopamine agonists and
cholinergic agents (Okuyama et al. 1987).
It is of interest that haloperidol
attenuated both the wet dog shakes and the
forepaw treading elicited by senktide. The
former behaviour is thought to be mediated by
5-HT, receptors (Arnt et al. 1984; Tricklebank
1985; Handley and Singh 1986) and haloperidol
has a low affinity for these sites. However,
alpha1-adrenergic mechanisms also appear to be
involved in 5-HT mediated wet dog shakes (Arnt
et al. 1984; Handley and Singh 1986) and
alpha1-adrenoceptor blockade may explain this
effect of haloperidol. Forepaw treading appears
to be mediated by 5-HT IA receptors (Tricklebank
et al. 1984; Tricklebank 1985) and haloperidol
is again inactive at these sites. However,
Tricklebank et al. (1984) found that forepaw
treading and other behaviours elicited by the
selective 5-HTLA agonist
8-hydroxy-2-(di-npropylamino)tetralin
(8-OH-DPAT) were blocked by haloperidol in
intact animals, although the effect of
haloperidol disappeared in reserpinized rats.
They did not have a good explanation for these
intriguing observations, and nor do we at this
time.
Our observations suggest that NK-3
tachykinin receptors may play a biologically
important role in the regulation of central
cholinergic and serotonergic neurons. It will be
of interest to further characterize these
interactions and examine their possible
relevance in neurodegenerative
disease.
-Banks RJ,
Mozley L, Dourish CT. The angiotensin
converting enzyme inhibitors captopril and
enalapril inhibit apomorphine-induced oral
stereotypy in the rat. Neuroscience.
1994;58(4):799-805
-Stoessl
AJ, Dourish CT, Iversen SD
Apomorphine-induced yawning in rats is abolished
by bilateral 6-hydroxydopamine lesions of the
substantia nigra Psychopharmacol; 1987; 93;
336-342
-Stoessl
AJ Effects of ageing on the behavioural
responses to dopamine agonists: decreased
yawning and locomotion, but increased stereotypy
Brain Research 1989; 495; 20-30
-Stoessl AJ,
Dourish CT, Iversen SD The NK-3 tachykinin
agonist senktide elicits yawning and chewing
mouth movements following subcutaneous
administration in the rat. Evidence for
cholinergic mediation. Psychopharmacology
(Berl). 1988; 95; 4; 502-506
-Cooper SJ;
Dourish CT Neural basis of drug induced
yawning in neurobiology of stereotyped behaviour
Oxford ed 1990
-Cooper SJ,
Rusk IN, Barber DJ. Yawning Induced by the
Selective Dopamine D2 Agonist N-0437 is Blocked
by the Selective Doparnine Autoreceptor
Antagonist (+)-UH 232. Physiol Behav.
1989;45:1263-1266
-Dourish
CT, SJ Cooper Yawning elicited by sytemic
and intrastriatal injection of piribedil and
apomorphine in the rat Pyschopharmacology 1985;
86; 175-181
-Dourish CT,
PH Hutson Bilateral lesions of the striatum
induced with 6-hydroxydopamine abolish
apomorphine-induced yawning in rats
Neuropharmacology 1985; 24; 11; 1051-1055
-Collins G, JM
Witkin et al Dopamine agonist-induced
yawning in rats: a dopamine d3 receptor mediated
behavior J Pharmacol Exp Ther 2005
Effects of
intrastriatal infusion of D2 receptor antisense
oligonucleotide
on
apomorphine-induced behaviors in the
rat.
Rajakumar N, Laurier L, Niznik HB, Stoessl
AJ.
Synapse. 1997;26(3):199-208.
Department of Clinical
Neurological Sciences, University of Western
Ontario, London, Canada.
An antisense oligonucleotide strategy was
employed to specifically deplete postsynaptic
striatal D2 receptors in order to determine the
possible role of presynaptic D2 autoreceptors in
mediating behavioral responses induced by low
doses of apomorphine. A
phosphorothioate-modified antisense
oligonucleotide complementary to the first 19
bases of the coding region of D2 receptor mRNA,
a scrambled sequence comprising the same bases,
or saline was infused bilaterally into the
striatum of adult rats, twice daily for 2 days
via indwelling cannulae. After an interval of
8-12 h, rats were habituated and challenged with
high (300 micrograms/kg; subcutaneous) or low
(50 micrograms/kg; s.c.) doses of apomorphine or
its vehicle (0.1% ascorbic acid). Yawning,
vacuous chewing mouth movements,
hypoexploration, and penile grooming induced by
low-dose apomorphine were unaffected by
antisense infusion into the striatum, whereas
stereotypic sniffing following high-dose
apomorphine was markedly suppressed.
Intrastriatal infusion of antisense resulted in
significantly diminished [3H]-raclopride
binding, while binding of [3H]-SCH 23390
(D1 receptors) and [3H]-WIN 35428
(dopamine transporter) was unchanged. D2 mRNA
levels determined by quantitative in situ
hybridization were normal in the striatum and
the substantia nigra. Our results confirm that
stereotypic sniffing is mediated via
postsynaptic D2 receptors in the striatum, and
favor the notion that behavioral responses
induced by low doses of apomorphine are mediated
by presynaptic D2 autoreceptors.
Behavioural effects
of selective tachykinin agonists in midbrain
dopamine regions.
Stoessl AJ, Szczutkowski E, Glenn B, Watson
I
Brain Res. 1991;565(2):254-62.
Department of Clinical
Neurological Sciences, University of Western
Ontario, London, Canada.
The effects of selective NK-1, NK-2 and NK-3
tachykinin agonists in midbrain dopamine cell
containing regions were investigated in the rat.
The NK-3 agonist senktide induced locomotion,
rearing and sniffing following infusion into the
substantia nigra pars compacta, and to a lesser
extent in the ventral tegmental area. These
behavioural responses were not seen following
infusion of the selective NK-1 agonist
[Sar9,Met (O2)11]SP or the NK-2 agonist
[N1e10]NKA4-10. In contrast, grooming
was induced only by the NK-1 agonist
administered into the substantia nigra. Yawning,
chewing mouth movements and wet dog shakes were
all seen following infusion of senktide into the
ventral tegmental area. These findings suggest
that (i) dopamine-mediated behavioural responses
seen following tachykinin administration into
the midbrain are dependent upon stimulation of
NK-3 tachykinin receptors, (ii)
tachykinin-induced grooming is mediated by
stimulation of NK-1 receptors and (iii) some of
the previously described 5-HT mediated
behaviours seen following administration of NK-3
tachykinin agonists are probably generated by
stimulation of 5-HT cell bodies in the ventral
tegmental area.