Yawning/cortical
activation induced by microinjection of
histamine into the paraventricular nucleus of
the rat
Yoshinari Seki, Ikuko Sato-Suzuki, Ichiro
Kita,
Mitsugu Oguri, Hideho Arita
Department of Physiology,
Toho University School of Medicine, Ohta-ku,
Tokyo Japan
-Kita
I, Kubota N, Yanagita S, Motoki C
Intracerebroventricular administration of
corticotropin-releasing factor antagonist
attenuates arousal response accompanied by
yawning behavior in rats. Neurosci.Letter
2008;433(3):205-208
-Kita
I, Yoshida Y, Nishino S. An activation of
parvocellular oxytocinergic neurons in the
paraventricular nucleus in oxytocin-induced
yawning and penile erection. Neurosci Res.
2006;54(4):269-275
-Kita I,
Sato-Suzuki et al.Yawning responses induced
by local hypoxia in the paraventricular nucleus
of the rat.Behavioural Brain Research
2000;117(1-2):119-126
-Kubota
N, Amemiya S, Motoki C, Otsuka T, Nishijima T,
Kita I. Corticotropin-releasing factor
antagonist reduces activation of noradrenalin
and serotonin neurons in the locus coeruleus and
dorsal raphe in the arousal response accompanied
by yawning behavior in rats. Neurosci Res.
2012;72(4):316-323
-Seki Y, Y
Nakatani, et al Light induces cortical
activation and yawning in rat Behav Brain Res
2003;140(1-2):65-73
-Seki Y,
Sato-Suzuki I, et al Yawning/cortical
activation induced by microinjection of
histamine into the paraventricular nucleus of
the rat. Behav Brain Res.
2002;134(1-2):75-82.
-Sato-Suzuki I,
Kita I, Oguri M, Arita H Stereotyped yawning
responses induced by electrical and chemical
stimulation of paraventricular nucleus of the
rat Journal of Neurophysiology,
1998;80(5)2765-2775
Introduction : We have reported that
a stereotyped yawning response can be evoked by
several chemical stimulations of the
paraventricular nucleus (PVN) in anesthetized,
spontaneously breathing rats. A typical yawning
response is induced by microinjection of
L-glutamate in the medial parvocellular
subdivision (mp) of the PVN, and is
characterized by an initial depressor response
and an arousal shift in electrocorticogram
(ECoG) followed by a single large inspiration
with mouth opening. Our concept that the PVN is
essential for the occurrence of yawning is
compatible with previous data of Argiolas and
Melis who demonstrated that lesions of the PVN
prevents yawning induced by apomorphine and
oxytocin in freely moving rats. On the other
hand, two physiological roles of the PVN
concerning yawning have been newly suggested by
our recent findings:
-1- An oxygen sensor may exist within the
PVN and yawning may be an arousal behavior
caused by higher brain ischemia, since
microinjection of cyanide into the mp caused the
stereotyped yawning response.
-2- The PVN may play an important role in
triggering arousal mechanisms, since
microinjection of orexin-A, a neuropeptide
involved in sleep and arousal mechanisms, into
the mp caused thestereotyped yawning
response.
Yawning is one typical symptom of motion
sickness. The involvement of histamine in neural
processes of motion sickness is suggested by
several pieces of evidence. In animal models of
motion sickness, the histamine level in the
hypothalamus increases after rotation. Another
piece of evidence comes from Takeda observation
that administration of fluoromethylhistidine, an
inhibitor of histamine synthesizing enzyme,
suppresses the signal of motion sickness in
rats. It is also clinically well known that
antihistaminics are effective in preventing
motion sickness in humans. These data on motion
sickness led us to speculate that the yawning
behavior might also be affected by histaminergic
neural transmission.
The neurons responsible for yawning are
proved to be the oxytocinergic parvocellular
neurons in the PVN projecting to the lower brain
stem. On the other hand, oxytocinergic
parvocellular neurons in the PVN are reported to
be activated by histamine. These two data
together support our idea that yawning might be
affected by histamine. The present study
focused on the yawning response induced by
microinjection of histamine into the PVN.
Attempts were further made to identify whether
the H1 receptor in the PVN mediates this
response. [...]
Discussion : We found that the
stereotyped yawning response was activated by
local application of histamine into the PVN,
which indicates that the PVN is a region
receiving histaminergic inputs from the
tuberomammillary nucleus (TM) of the posterior
hypothalamus, a region where histaminergic
neurons are densely located. Our view that the
PVN is a histamine sensitive site is supported
by three pieces of evidence.
-1- Panula demonstrated that histaminergic
neurons originating in the TM project to other
brain areas including the PVN. Another piece of
evidence comes from an autoradiographic study
showing the presence of histamine receptor in
the PVN. Finally, Bealer and Abell caused
effective cardiovascular responses by applying
histamine through microdialysis probes into the
PVN. Our data showing that histamine
administered into the PVN induced yawning,
together with an arousal shift in the ECoG,
confirms the results of previous studies which
showed that histamine is an important
neurotransmitter in arousal regulation.
Experimental lesioning of the posterior
hypothalamus has been reported to induce a state
of somnolence in rats . In addition, the
decrease in histamine content of the brain
suppresses wakefulness in rats. Conversely,
histaminergic drugs cause wakefulness in rats.
These arousal effects of histaminergic systems
are thought to be mediated by H1 receptors. It
is also clinically well known that
antihistaminics induce drowsiness. Moreover,
particularly high levels of histamine receptors
are present in area involved in arousal, i.e.
the cerebral cortex, cholinergic cell groups in
the mesopontine tegmentum (MPT) and in the basal
forebrain, as well as the locus
coeruleus (LC).
-2- We further indicate that not only
histamine but also PVN is involved in the
arousal signaling pathway, since microinjection
of histamine into the PVN induced the arousal
shift in the ECoG together with the yawning
response. It should be emphasized that no other
area in or around the PVN elicited the
arousal/yawning response. These data may
strengthen the hypothesis proposed in our
previous study, that the PVN is a site involved
in the arousal mechanism. According to the
arousal pathway referred to as histaminergic
transmission, several possible pathways are
nominated. Arousal may be due to histaminergic
neurons originating in the TM directly
projecting to the cerebral cortex. On the other
hand, it may be due to histaminergic neurons
projecting to the cholinergic cell groups in the
MPT or in the basal forebrain as well as the LC.
Here we suggest another signaling pathway of the
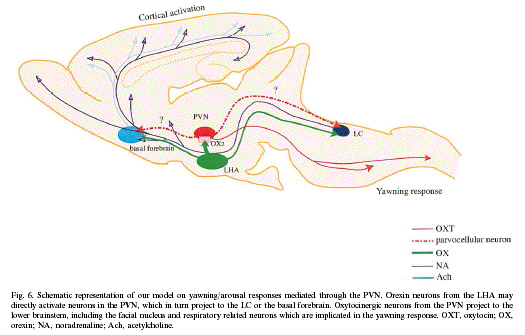
arousal mechanism via the PVN. Namely,
histaminergic neurons from the TM may directly
activate parvocellular neurons in the PVN, which
in turn project to the basal forebrain or the
LC. We previously reported that the neurons
responsible for yawning might be the
oxytocinergic parvocellular neurons in the PVN
projecting to the lower brainstem. Our
suggestion was mainly based on the report of
Sawchenko and Swanson who demonstrated that
oxytocinergic parvocellular neurons in the PVN
send descending axons to the lower brain stem
which region is involved in arousal,
respiratory, cardiovascular, and other autonomic
functions. According to this notion, the
present data can be explained by the possibility
that histamine activates the oxytocinergic
parvocellular neurons in the PVN projecting to
the lower brain stem. This possibility is
strongly supported by the data of Kjær who
reported that c-fos expression in oxytocinergic
parvocellular neurons within the PVN is induced
by central administration of histamine. They
further suggested that histamine might be
indirectly involved in the regulation of neurons
in the lower brain stem and the spinal cord via
the PVN.
-3- Although we suggest that the
oxytocinergic parvocellular neurons in the PVN
are involved in yawning, we can not exclude
the possibility that the PVN may contain more
than one pathway mediating this response. For
instance, ACTH injected into the PVN and
surrounding periventricular region induces
yawning that is not prevented by oxytocin
antagonist. Corticotropin releasing factor (CRF)
neurons within the PVN could be responsible for
the ACTH-induced yawning, however, there is
little evidence that CRF neurons project to the
lower brain stem which region is involved in
arousal, respiratory, cardiovascular, and other
autonomic functions regarded to yawning.
We indicate that histamine activates the H1
receptor in the PVN, since the yawning response
induced by histamine was inhibited by
pretreatment with pyrilamine, an H1 receptor
antagonist. Moreover, microinjection of HTMT
dimaleate, an H1 receptor agonist, into the PVN,
evoked the yawning response. Similarly to our
data, Gower reported that H1 receptor antagonist
injected subcutaneously into conscious rats
inhibited the apomorphine-induced yawning
response. With regard to this view, an
autoradiographic study revealed the presence of
H1 receptor in the PVN. The existence of H1
receptor in the PVN has also been examined in
functional studies.
For example, Bealer demonstrated that
histamine evokes norepinephrine as well as
oxytocin release by activation of H1 receptors
in the PVN. H1 receptors in the PVN are also
reported to be involved in histaminergic
suppression of food intake. Although H1 receptor
may be important in the modification of the
yawning/arousal response, the role of other
types of histamine receptors cannot be fully
eliminated. In this connection, Ferrari and
Baggio reported that H2 receptor antagonist
dose-dependently antagonized the yawning and
penile erection induced by apomorphine.
In line with their data, yawning response
was sometimes induced by H2 receptor agonist
into the PVN in the present study. However, H2
receptor agonist induced a yawning response with
a much longer onset latency compared with that
of histamine or H1 agonist. In addition,
pretreatment with H2 receptor antagonist did not
block the yawning response induced by histamine
in our study. Further study is required before
the precise mechanism of histamine actions on
yawning/arousal responses can be
identified.
We noticed that a pressor response appeared
after the yawning behavior induced by
microinjection of histamine into the PVN, which
lasted over 90 s. In this connection, Bealer and
Abell demonstrated that administration of
histamine through microdialysis probes in the
PVN increased BP in conscious rats. It is
suggested that the PVN is a brain site where
histaminergic and noradrenergic systems interact
to regulate BP, and the pressor response evoked
by histamine may be due to adrenoreceptor
stimulation of vasopressin release. Although the
increase in HR was prominent after injection of
H2 agonist into the PVN (data not shown), we
could not observe a clear increase in HR by
histamine injection into the PVN, which result
differs from that in conscious animals. The lack
of changes in HR by histamine in this study
might be attributed to the possibility of
anesthetics used, since anesthesia is well known
to affect the cardiovascular and autonomic
responses.
Since yawning is one typical symptom of
motion sickness a disorder which is modified by
histaminergic transmission, the present results
can be explained by the notion that yawning
is triggered by motion sickness which signaling
pathway is mediated by histaminergic
neurons. According to our data showing that
yawning was inhibited by H1 receptor antagonist,
together with previous data showing that motion
sickness is suppressed by H1 antagonist, it is
likely that histamine induces yawning as well as
motion sickness through H1 receptor within the
PVN.
-Kita
I, Yoshida Y, Nishino S. An activation of
parvocellular oxytocinergic neurons in the
paraventricular nucleus in oxytocin-induced
yawning and penile erection. Neurosci Res.
2006
-Kita I,
Sato-Suzuki et al.Yawning responses induced
by local hypoxia in the paraventricular nucleus
of the rat.Behavioural Brain Research 2000; 117;
1-2; 119 - 126
-Sato-Suzuki I,
Kita I; Oguri M, Arita H Stereotyped yawning
responses induced by electrical and chemical
stimulation of paraventricular nucleus of the
rat Journal of Neurophysiology, 1998; 80, 5;
2765-2775
-Sato-Suzuki I,
I Kita, YSeki, M Oguri, H Arita Cortical
arousal induced by microinjection of orexins
into the paraventricular nucleus of the rat
Behavioural Brain Research 2002; 128;
169-177
-Seki Y, Y
Nakatani, et al Light induces cortical
activation and yawning in rat Behav Brain Res
2003; 140; 1-2; 65-73
-Seki Y,
Sato-Suzuki I, et al Yawning/cortical
activation induced by microinjection of
histamine into the paraventricular nucleus of
the rat. Behav Brain Res.
2002;134(1-2):75-82.