Introduction : Current knowledge on
yawning behavior in Old World monkeys can be
summarized as follows. Three types of yawns
have been distinguished: physiological yawn,
emitted during periods of transition from waking
to sleeping, or during the downward phases of
cyclic arousal (Deputte
1978); stress yawn, occurring in conflict or
anxiety-producing situations (Hinde & Rowell
1962); and threat yawn, involving canine display
and characterized by direct orientation toward
another animal (Hall & Devore1965). Yawning
is displayed more frequently by males than
females (e.g. Redican 1975). Its rate of
occurrence is influenced by plasma levels of
androgens; injection of testosterone propionate
in adult rhesus females induced a 20-fold
increase in yawning behavior (Goy & Resko
1972). Male yawning increases through
adolescence and jumps dramatically as males
enter early adulthood, whereas female yawning
increases only slightly, if at all, after
infancy (Hadidan
1980). The highest-ranking male in the dominance
hierarchy tends to yawn at a much higher rate
than other group members. Adult male macaques
appear able to produce yawning voluntarily
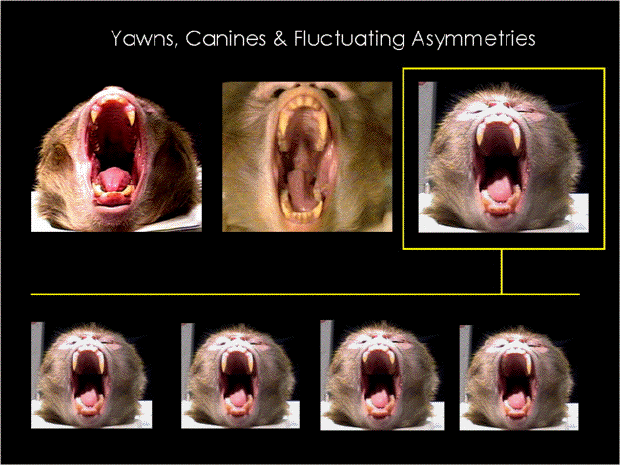
(Anderson & Wunderlich 1988). See photos in
the bottom.
Most of the above information comes from
either anecdotal reports or laboratory studies
involving small numbers of subjects. Among
published papers, only one naturalistic study
has quantitatively investigated the spontaneous
occurrence of yawning in a social group of
nonhuman primates (Hadidan
1980). Surprisingly, the ethology of yawning bas
been studied more systematically in humans than
in nonhuman primates (e.g Baenninger
1987; Provine
et al. 1987; Schino
& Aureli 1989).
Adopting a naturalistic and quantitative
approach, the present study aimed at documenting
the occurrence of yawning behavior in two
species of macaques: Macaca fascicularis and M.
fuscata. In particular, the influence of age,
sex, and dominance rank on yawning was
investigated. Differences in the frequency of
yawning by individuals from different age-sex
classes within the group were analyzed with a
view to elucidating proximate causation of
yawning. Because of the methodological
difficulties of distinguishing the various types
of yawns according to their functional
significance and context of occurrence, the
present study deals only with the apparent
structural pattern and not its functional types.
[...]
Discussion : The two macaque groups
of this study showed major differences in the
overall frequency of yawning and in the
influence that sex, age, and dominance rank
exerted on yawning behavior. Japanese macaques
yawned much more than long-tailed macaques.
Although the long-tailed macaque data may
underestimate the frequency of yawning, it seems
unlikely that methodological differences could
account for the 10-fold difference in yawning
rates between the two species. In the
long-tailed macaques, sex differences in
frequency of yawning only emerged after sexual
maturity; yawning rates increased significantly
in both males and females as they reached sexual
maturity; and, among males, rank was positively
correlated with frequency of yawning.
Differently, in the japanese macaques, males,
both mature and immature, yawned more than
same-aged females; sexual maturity was
associated with an increase in yawning in males
only; and male rank did not correlate with
frequency of yawning.
Even though more than one group would need to
be examined to advance hypotheses concerning
interspecific distinctions, it may be
interesting to discuss the differences between
the two study groups in the light of comparative
data on the social structure of macaque species.
Caldecott (1986) has proposed a two group
classification of macaque societies. Species in
the first group (M. mulatta, M. fuscata, M.
nemestrina, and M. silenus) are characterized by
antagonistic inter-male relationships and a
multiple-mount copulation pattern. Species in
the second group (M. sylvanus, M. radiata, and,
based on recent data by Thierry (1985) and
DeWaal (1989), M. arctoïdes and M.
tonkeana) show relaxed internale relationships
and a single-mount copulation pattern. Based on
the inconsistent copulatory pattern of M.
fascicularis (both single- and multiple-mount
copulations), considers this species as
intermediate between the two groups. Comparative
data on agonistic behavior support the view that
the long-talled macaque occupies an intermediate
position between the two groups of Caldecott's
classification. Taken together, these data
suggest that social relationships in M. fuscata
are more tense than those found in M.
fasciculars. It is possible that the
interspecific difference in yawning rates that
we found in this study reflects such a
difference in the social structure of the two
species.
Regardless of interspecific differences, the
overall results of this study support only in
part the finding that yawning is largely
influenced by plasma concentrations of androgens
(Phoenix & Chambers 1986). On the one hand,
we found that mature males, the individuals with
the highest concentrations of androgens, yawned
much more than any other age-sex class, and that
male yawning increased dramatically after sexual
maturity, when the plasma levels of testosterone
start to rise. Also the finding that, after the
attainment of sexual maturity, long-tailed
macaque females showed an increase in frequency
of yawning may accord with an etiological
explanation implicating an activational role of
androgens: sexually-mature female monkeys
secrete androgens from the ovary as well as from
the adrenal gland (Hess & Resko 1973).
On the other hand, the increase in female
yawning after sexual maturity was recorded only
in the long-tailed macaque group. In addition,
in the Japanese macaque group, sex differences
in frequency of yawning were clearly evident
prior to the attainment of sexual maturity,
indicating that the higher rate of male yawning
was not simply due to plasma concentrations of
androgens. Although this latter finding can be
explained by hypothesizing an organizational
role for androgens (ie, in the prenatal period,
the male hormone would organize and
differentiate the neural tissues that mediate
the display of yawning), these results suggest
that social factors are also important in
influencing the age-sex class distribution of
yawning behavior. For example, it is possible
that, among immature Japanese macaques, yawns
were emitted more often by males than by females
because the males were generally more tense, a
possible result of their more frequent
involvement in agonistic behavior.
The role of social factors clearly emerges
from the data on the relationship between male
dominance rank and yawning. The finding that
higher-ranking long-tailed macaque males yawned
more than subordinate males fits well with the
notion that yawning is a mild form of threat.
Yet, this was not true for the Japanese macaque
males. This result brings to attention the
hypothesized dual function of emotional yawning
in Old World monkeys (Redican 1982). It is
possible that most of the yawns emitted by
low-ranking males were a result of conflict or
stress, whereas yawns by high-ranking males were
made as intimidating displays. If this was the
case, the same rate of yawning by males of
different rank shoul d be viewed as the final
outcome of two different motivational processes:
intention of eliciting submission from another
animal (threat yawn) and response to stressful
circumstances (stress yawn). The difficulty of
testing this hypothesis relates to the general
problem of distinguishing between primate
displacement activities (e.g. scratching,
self-grooming, yawning, and shaking) and the
same behavior patterns carried out in a "normal"
context, ie. as comfort behaviors (Diezinger
& Anderson 1986).
In conclusion, the present study shows
that macaque yawning has a complex causation
which is likely to result from an interaction
between hormonal and social factors.
However, the heterogeneity of the few available
data, including those in the present study,
indicates that there is a need for further
naturalistic studies of yawning behavior in
different primate groups and species.
BAENNINGER,
R. 1987: Some comparative aspects of yawning in
Betta splendens, Homo sapiens, Panthera leo, and
Papio sphinx. J. Comp. Psychol. 101,
349-354
CALDECOTT, J. 0. 1986: Mating patterns,
societies, and the ecogeography of macaques.
Anim. Behav. 34, 208-220.
DEPUTTE, B.
1978; Etude du bâillement chez deux
espèces de Cercopithecidae, Cercocebus
albigena Gray et Macaca fascicularis Raffles:
Recherche des facteurs de causalité et de
fonction. Mise en évidence des facteurs
socio-bioénergétiques. Thesis,
Université de Rennes
DE WAAL, F. B. M. & LUTTRELL, L. M.
1989: Toward a comparative socioecology of the
genus Macaca: different dominance styles in
rhesus and stumptail monkeys. Am. J. Primatol.
19, 93-109
DIEZINGER, F. & ANDERSON, J. R. 1986:
Starting from scratch: a first look at a
"displacement activity" in group-living rhesus
monkeys. Am. J. Primatol. 11, 117-124.
Goy,
R. W. & RESKO, G. A. 1972: Gonadal
hormones and behavior of normal and
pseudohermaphroditic nonhuman female primates.
Rec. Progr. Horm. Res. 28, 707-733
HADIDIAN, J.
1980: Yawning in an Old World monkey, Macaca
nigra (Primates: Cercopithecidae) Behaviour 75,
133-147
HALL, K. R. L. & DEVORE, 1. 1965: Baboon
social behavior. In: Primate Behavior: Field
Studies of Monkeys and Apes. (DEVORE, I., ed.)
Holt, Rinehart & Winston, New York, pp.
53-110.
HESS, D. L. & RESKO, G. A. 1973: The
effects of progesterone on the patterns of the
testosterone and estradiol concentrations in the
systemic plasma of the fernale rhesus monkey
during the intermenstrual period. Endocrinology
92, 446-453
HINDE, R. A. & ROWELL, T. E. 1962:
Communication by posture and facial expression
in the rhesus monkey. Proc. Zool. Soc. Lond.
138, 1-21.
MARTIN, P. & BATESON, P. 1986: Measuring
Behaviour. Cambridge Univ. Press,
Cambridge.
NAPIER, P. H.
1967: A Handbook of Living Primates. Acad.
Press, London.
PHOENIX,
C. H. & CHAMBERS, K. C., 1986: Threshold
for behavioral responses to testosterone in old
castrated male rhesus macaques. Biol. Reprod.
35, 918-926.
PROVINE, R.
R., HAMERINK, H. B. & CURCHACK, B. C. 1987:
Yawning: relation to sleeping and stretching in
humans. Ethology 76, 152-160.
REDICAN, W. K. 1975: Facial expressions in
nonhuman primates. In: Primate Behavior:
Developments in Field and Laboratory Research,
Vol. 4. (RoSENBLum, L. A., ed.) Acad. Press, New
York, pp. 103-194.
1982: An evolutionary perspective on human
facial displays. In: Emotion in the Human Face,
2nd ed. (EKMAN, P., ed.) Cambridge Univ. Press,
Cambridge, pp. 212-280.
SCHINO,
G. & AURELI, F. 1989: Do men yawn more
than women? Ethol. Sociobiol. 10, 375-378.
THIERRY, B. 1985: Patterns of agonistic
interactions in three species of macaques (M.
mulatta, M. fascicularis, M. tonkeana). Aggress.
Behav. 11, 223-33
« It is
ironic that testosterone "the male sex hormone,"
is more closely associated with the yawning rate
than with the mounting or intromitting rates
» Charles Phoenix
Sexual
steroids
exert several effects on both central
dopaminergic and oxytocinergic systems by acting
either at the genomic or membrane level