Earlier studies have demonstrated that
interesting and important circulatory changes
accompany excessively prolonged or severe
expiratory activities such as crying, coughing
and straining. Circulatory changes would also be
expected to accompany excessively prolonged or
severe inspiratory acts such as dying gasps,
yawns and sighs. Yet, none are described in the
current literature and textbooks. It is
interesting that Stephen Hales in 1733 observed
that deep sighing and respiratory efforts of
dying mares increased systematic arterial
pressure. He believed the greater motion of the
lungs caused the blood to pass more freely and
in greater quantity to the left heart.
The observations reported in the present
study are limited principally to the effects of
normal respiration, deep breathing and dying
gasps, since yawns and sighs could not be
induced experimentally in dogs. However, the
data have allowed appreciation of the
hemodynamic changes induced by sighs and
yawns.
METHODS. Morphine sulfate, 5 mg per
kg, was administered subcutaneously 30 minutes
prior to the experiments. All operative
procedures were accomplished with the aid of
local infiltrations of 1 per cent procaine
hydrochloride. In those experiments where it was
necessary to enter the chest, the appropriate
intercostal nerves were blocked.
Changes in the pressure relationships
between the thoracic and abdominal cavities were
measured since they modify venous return from
the abdominal reservoirs to the right heart.
Balloons fastened to leaden tubes and inserted
into the abdominal and thoracic cavities were
connected to a differential manometer so that
the intrathoracic pressure was subtracted from
the abdominal pressure. This differential
pressure has been called the abdominal thoracic
pressure gradient. Changes in this gradient
modify venous return to the right heart and can
result from unequal increases or decreases in
the intrathoracic or intra-abdominal pressures.
When it was unnecessary to enter the chest, the
balloons were inserted through the mouth and
into the esophagus or stomach. Long balloons
only partially filled were used since they tend
to dissipate pressure changes from the activity
of the alimentary canal but register the changes
transmitted to them from the intrapleural space
and abdominal cavity. These esophageal and
gastric pressures were considered to change in
parallel with thoracic and abdominal
pressures.
Hollow sounds constructed from stainless
steel tubing G 12 to 16 and adapted to the size
of the animal were connected to other optical
manometers and were inserted 1, down the left
carotid and into the left ventricle, and 2, down
the right external jugular and into the right
ventricle. Saline infused through these sounds
at the rate of about 1 cc. per minute reduced
the danger of obstruction by coagulation. This
made it possible to obtain continuous
simultaneous pressure records from the left and
right ventricles in the closed chest. Both gross
and net ventricular pressures were obtained. The
gross pressures were obtained with simple
manometers (1), as commonly measured from the
ventricular cavities and show both active and
passive changes. The net ventricular pressures
are the gross pressures minus the intrathoracic
pressure and were obtained with differential
manometers (4) where the ventricular pressures
were led to the rear chambers and the
intrathoracic pressure was led to the front
chambers of the manometers in the usual manner.
These net ventricular pressures show only active
changes and are the pressures actually
distending the heart cavities. They are the
effective blood pressures to the blood vessels
and organs within the thorax. The net left
ventricular systolic pressure can be considered
equivalent to the net coronary systolic
pressure. The net right ventricular systolic
pressure be can considered equivalent to the net
pulmonary arterial systolic pressure. In
addition to the net coronary systolic and the
net pulmonary arterial systolic pressures, when
necessary to determine net pulmonary venous and
net inferior vena cava pressures, they can be
obtained from such records by considering
thefollowing as equivalents: the net right
ventricular diastolic pressure is equivalent to
the net inferior vena cava pressure; the net
left ventricular diastolic pressure is
equivalent to the net pulmonary venous
pressure.
Similar methods supplied measurements of the
gross and net systemic arterial pressure to the
central nervous system. The gross systemic
arterial pressure acts to push blood to organs
including the brain and spinal cord. The
pressure within the craniospinal cavity acts to
hinder blood inflow. Therefore, the effective
arterial pressure to the central nervous system
is the arterial pressure minus the cerebrospinal
fluid pressure. This effective pressure has been
called the net arterial pressure to the brain
and spinal cord and has been calculated from
simultaneous measurements of the arterial
pressures and cerebrospinal fluid pressures. The
cerebrospinal fluid pressure was recorded from
no. 18 or no. 20 G needles inserted in three
animals into the subarachnoid space in the
region of the first or second lumbar
vertebra.
Acute cardiac arrest was produced by
electrically induced ventricular fibrillation.
In four dogs the electrodes were placed upon the
ventricles while inserting the thoracic
balloons. The chest was then closed and the
pneumpthorax was reduced. In two other animals
where the chest was not opened an insulated
sound with the end serving as the stimulating
electrode was introduced down the external
jugular into the right ventricle. An indifferent
electrode was placed upon the skin over the
heart. The various gross and net pressures were
then recorded while stimuli were applied to the
ventricle using two batteries and a Harvard
Inductorium at full strength. (...)
RESULTS AND DISCUSSION. First the
effects of normal respiration will be described.
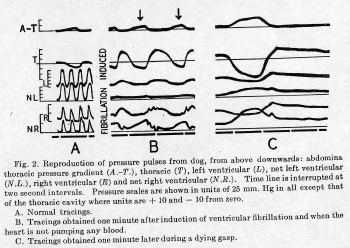
As shown in figure 2A, normal inspiration
(arrow) lowers the thoracic pressure and
increases the abdominal thoracic pressure
gradient which increases venous return td the
heart from the portal system and the inferior
vena cava. During inspiration the net right
ventricular pressure pulses show a higher pulse
pressure and a peak with a wider plateau, see
second and third NR pressure pulses of figure
2A. These changes indicate a larger and more
prolonged effective ejection period without any
appreciable change in the duration of systole.
These changes are interpreted as characteristic
of an increased right cardiac output secondary
to an increased filling with inspiration.
When ventricular fibrillation was induced
with strong tetanizing electrical stimuli
applied directly to the heart in the closed
chest, respiratory activity continued for
several minutes after effective cardiac action
had ceased. The deep foreeful natural breathing
(see fig. 2B) produced significant changes in
the blood pressure and circulated blood. During
inspirations (arrows) the abdominal thoracic
pressure gradient is increased; the gross right
ventricular pressures returned toward zero; but
the effective net right ventricular pressure
(NR) increased. As shown in figure 2B, it
reached values between 25 and 30 mmHg which were
approximately equal to those produced by the
right ventricle when it was contracting (fig.
2A). Simultaneously these deep respiratory
efforts produced only small changes in the
effective net left ventricular pressures (NL).
It can be concluded from these data that
repeated deep forceful breathing, even in the
presence of complete cardiac arrest, repeatedly
increased venous return to the right ventricle
and to the pulmonary vessels while moving a
smaller quantity of blood from the pulmonary
vessels on into the left ventricle. This also
means that deep forceful breathing in the
presence of cardiac failure can contribute to
pulmonary engorgement by transferring blood from
the abdominal venous reservoirs to the pulmonary
vessels.
Since sighs and yawns are modified deep
breaths they also should increase venous return
to the right heart. It has been observed by one
of the authors that small children when asleep
will usually yawn if they are lifted into an
upright position. Sometimes the yawn would occur
repeatedly each time they were changed to an
upright position. This yawn could be
physiologically required to increase air
exchange or to increase venous return. There is
no reason to expect any real need of increased
air exchange. However, it is conceivable that a
sudden change to an upright position in an
individual whose muscles are relaxed would
retard venous return sufficiently so that a
mechanism to increase venous return
operates.
A yawn to be truly satisfying frequently is
accompanied by a stretch. When present the
stretch not only contributes to venous return
but also, like the dying gasp, tends to divert
blood to the heart and central nervous
system.
Circulatory changes are pronounced during
dying gasps. The effective net
right ventricular pressure is elevated to
values as high as 50 mmHg (see fig. 2C). The
pulmonary vessels become so engorged that blood
is pushed through to the left ventricle and
elevates the effective net pressure in that
chamber to 30 to 50 mmHg.
These dying gasps not only increase venous
return but they cause significant coronary blood
flow. Blood is pushed or diverted into the
coronary vessels as a result of the extensive
skeletal muscle activity and the elevated
abdominal pressure, both of which squeeze blood
vessels and increase peripheral resistance.
Simultaneously the intrathoracic pressure is
reduced so that peripheral resistance in the
coronary vessels is lowered while the effective
net coronary arterial pressure is elevated.
In similar experiments net cerebrospinal
arterial pressures of 20 to 40 mmHg were
produced by dying gasps. Therefore blood flow to
the central nervous system is also accomplished
by dying gasps. The mechanisms responsible for
blood flow to the central nervous system were
only slightly less effective than those
described above which produced coronary flow,
since dying gasps lower the cerebrospinal fluid
pressure only slightly less than they lower the
intrathoracic pressure. This is due to the fact
that the cerebrospinal fluid pressure is
influenced by the algebraic sum of the
intrathoracic and abdominal pressure, though
unpublished data indicate that it follows the
intrathoracic pressure more closely than it
follows the abdominal pressure.
These gasps produce air exchange and move
blood to the vital areas, whereas mechanical
resuscitators provide air exchange but do not
cause any significant blood flow to the vital
areas.
Cases have been encountered where patients
apparently dead and without any evidence of
heart action have again developed palpable pulse
and audible heart contractions immediately after
making dying gasps. Similar observations have
probably been passed off with the remark "I must
have been wrong when I failed to observe any
pulse the first time." it is conceivable however
that the blood pumping action of the dying gasps
contributed to the return of effective cardiac
contractions.
SUMMARY AND CONCLUSIONS
Gross and net, left and right ventricular
pressures are recorded from dogs without
operative entrance into the chest by means of
hollow sounds inserted down the left carotid
into the left ventricle and down the right
jugular into the right ventricle.
Normal inspiration increases venous return
to the right heart and produces contour changes
characteristic of larger and more prolonged
effective ejection without significantly
changing the duration of systole.
Dying gasps, deep breathing, yawns and sighs
which are generally considered as respiratory
acts, markedly increase venous return. In the
presence of cardiac arrest, dying gasps pump
blood through the lungs and temporarily provide
blood flow to the vital areas, the central
nervous system and heart. Effective net pressure
as great as 50 mmHg in the pulmonary artery, 50
mmHg in the coronary arteries and 40 mmHg in the
central nervous system arteries were created by
dying gasps in dogs where cardiac action had
ceased.
-Legendre
R, Piéron H.
De la propriété
hypnotoxique des humeurs
développée au cours d'une veille
prolongée C.R. Société de
Biologie de Paris 1912; 70; 210-212
-Bouyssou M,
Tricoire J Experimental detection of a
cervical arousal mechanism of yawning, enhancing
intracerebral (CSF) fluid pressure J Dental Res
1985; 64; 721
-Lepp
FH Remarques sur la signification
physiologique du bâillement Bull Group Int
Rech Sci Stomtol Odontol 1982; 25; 251-290
-Nolman
B Yawning, cerebral fluid and the lymphatic
pump 2006
-Oreskovic D;
Klarica M; Vukic M The formation and
circulation of cerebrospinal fluid inside the
cat brain ventricles : a fact or an illusion ?
Neuroscience letters 2002; 327; 103-106
-Patra P, Gunness
TK, Robert R Physiologic variations of the
internal jugular vein surface, role of the
omohyoid muscle, a preliminary echographic study
Surg Radiol Anat 1988; 10; 2; 107-12
-Schniter E The
evolution of yawning : why do we yawn and why is
it contagious ? thèse 2001
-Schroth G, Klose U
Cerebrospinal fluid flow; Physiology of
respiration-related pulsations. Neuroradiology
1992; 35; 1; 10-15
-Walusinski
O Prostaglandines, adénosine, sommeil
& bâillements 2004
-Woodbury R, B
Abretj Influence of dying gasps, yawns and
sighs on blood pressure and blood flow Am J
Physiol 1944; 142; 721-726