-
- Pourquoi
bâillons-nous ? Revue des théories
d'Hippocrate à nos jours Walusinski O
2014
-
- Abstract : The temporomandibular
joint is one of the most taxed joints of the
body. 5% of the population feel pain when
opening the mouth wide and 3% while chewing and
yawning, during which jaw opening reaches its
maximal course. As testifying during a
pandiculation, a close functional coupling
between the temporomandibular and the
cranio-cervical regions is triggered by the
brainstem through trigeminal somatosensory
projections onto the motoneurones of the
masseters and neck muscles during
jaw-opening/closing, and notably during yawning,
like a "trigemino-neck reflex". The neural
networks, required for these innate activities,
share phylogenical, ontogenical and anatomical
links with the brainstem reticular formation
activated during wakefulness and REM sleep. By
these links, yawning appears as a behavior
(fixed action pattern inverse of REM sleep)
which procures an arousal reinforcement through
the powerful stretch and the neuromuscular
rewiring induced. Excessive mouth opening while
yawning is the commonest cause of
temporomandibular joint dislocation. Pain
elicited by yawning is a well-recognized
clinical phenomenon in patients with cranial
neuralgia or dystonia, temporomandibular joint
dysfunction syndrome or Eagle syndrome.
-
- Résumé : L'articulation
temporo-mandibulaire est une des articulations
les plus sollicitées de l'organisme. 5%
de la population éprouve des douleurs
à l'ouverture de bouche et 3% notamment
en mâchant et en bâillant. En effet,
c'est au cours du bâillement que
l'ouverture de bouche et du pharyngo-larynx est
maximale. Comme le montre une pandiculation, il
existe un lien fonctionnel étroit,
régulé par le tronc
cérébral, entre les articulations
temporo-mandibulaires et le rachis cervical,
véritable "réflexe
trijémino-nuqual", pendant lequel les
projections proprioceptives du trijumeau
stimulent les motoneurones des masséters
et de la musculature cervicale. Ce réseau
neuronal, nécessaire à ces
comportements innés, partage des liens
phylogéniques, ontogéniques et
anatomiques avec la formation
réticulée activatrice du tronc
cérébral participant à
l'éveil et au sommeil paradoxal. Par ces
liens, le bâillement apparaît comme
un comportement (action coordonnée,
innée, inversant les effets hypotonisant
du sommeil paradoxal) qui procure une
stimulation de l'éveil par les influx
proprioceptifs en retour de la très
puissante contraction musculaire
engendrée. L'extême ouverture de la
bouche au cours du bâillement est la cause
la plus fréquente de la luxation de la
mâchoire. Les douleurs
déclenchées par le
bâillement sont cliniquement facilement
identifiables chez des patients souffrant de
dystonie ou d'algies de la face, de
désordres temporo-mandibuaires ou d'un
syndrome d'Eagle.
-
Au sein de la population adulte, 5% des
personnes interrogées se plaignent de
douleurs lors de la complète ouverture de
la bouche et 3% en particulier au cours du
bâillement (Heloë, 1979). Les
articulations temporo-mandibulaires sont parmi
les articulations les plus sollicitées de
l'organisme. Elles participent au langage,
à la mastication mais aussi à de
multiples expressions émotionnelles du
visage comme le rire, la peur, la honte, la
colère, etc (Duchenne de Boulogne, 1862).
Un comportement, le bâillement, a lui,
pour caractéristique, de leur imposer
leurs amplitudes extrêmes: ouverture
maximale, suivie d'une occlusion complète
de la bouche qui n'existe dans aucune autre
situation physiologique.
-
- Appareil manducateur, cou et
bâillement.
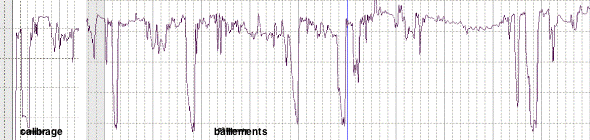
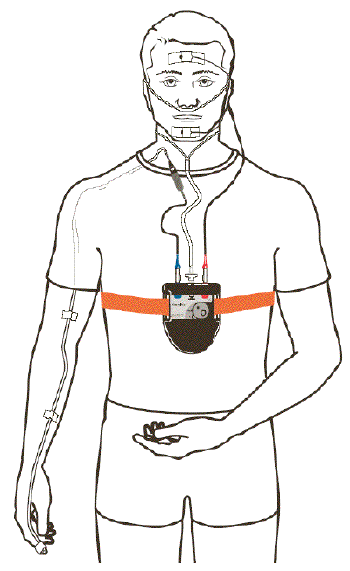
- Un appareil de dépistage ambulatoire
du syndrome des apnées du sommeil
(Somnolter©) recherche les épisodes
de ronflements et d'efforts ventilatoires par la
mesure, de manière continue, des
mouvements de la mandibule et par l'estimation
de la distance intermaxillaire. Comme l'indique
la figure 1, l'amplitude d'ouverture de la
bouche lors d'un bâillement
spontané est plus importante que
l'ouverture volontaire maximale de la bouche
lors du calibrage de l'appareil. A noter que
l'occlusion qui suit la fin du bâillement
apparaît également plus
accentuée que l'occlusion volontaire.
L'analyse détaillée d'un
bâillement permet d'en comprendre les
mécanismes.
-
 -
- Les mouvements de la tête font partie
intégrante du cycle ouverture/fermeture
de la bouche nécessaire à la
mastication, à l'élocution, au
chant, mais aussi au bâillement (Abrahams,
1988, 1993). D'un point de vue
phylogénétique, chez toutes les
espèces, ce couplage fonctionnel a une
valeur adaptative, sélectionnée,
car elle assure une meilleure capacité
à saisir des proies mais aussi à
se défendre et à combattre. Les
afférences sensori-motrices oro-faciales
(trijumeau) sont nécessaires au
contrôle de la motricité
cervico-céphalique, c'est à dire
que l'activité fonctionnelle mandibulaire
est une association de mouvements
synchronisés de la nuque et de la
mâchoire dans lesquels interviennent les
articulations temporo-maxillaires, le rachis
cervical et sa musculature (Zafar, 2000).
L'action des muscles masséters et
cervicaux est synchronisée par une
commande motrice commune automatique
générée par le tronc
cérébral, ("Central pattern
generating circuits" Marder, 2005) c'est
à dire les paires crâniennes V,
VII, IX, X, XI, XII. Toutes ces structures
motrices sont activées pendant les
bâillements. T. Humphrey (1968) a
parlé d'un réflexe
trijémino-nuqual.
-
- On appelle une pandiculation une ouverture
maximale de la bouche associée à
une contraction généralisée
des muscles antigravitaires ou antigravifiques
(André-Thomas, 1949) des quatre membres
avec extension complète de l'ensemble du
rachis, portant en arrière la tête.
Ce puissant étirement s'accompagne d'une
activation de tous les muscles de la face, et
très curieusement, d'une contraction
simultanée des muscles ouvrant la bouche
(mylo-hoïdiens et digastriques) et de ceux
assurant sa fermeture
(ptérygoïdiens, masséters,
temporaux), expliquant les contraintes
importantes subies alors par les articulations
temporo-mandibulaires (Forte, 1982).
Simultanément, le plus grand muscle de
l'organisme, le diaphragme, se contracte
massivement provoquant une ample inspiration par
des voies aériennes supérieures
ouvertes à l'extrême. J. Barbizet
(1958) a montré que le diamètre du
pharyngo-larynx est multiplié par quatre
par rapport à la position de repos. Le
bâillement apparaît ainsi comme une
pandiculation partielle, c'est à dire
sans contraction des membres et du tronc et rend
compte de la synchronisation neuro-musculaire
bucco-cervicale. La composante ventilatoire du
bâillement et de la pandiculation ne sont
donc que la conséquence d'une
activité musculaire et non le primum
movens. Ceci explique l'absence de toute
modification des gaz du sang pendant et
après le bâillement et infirme la
notion ancienne d'une oxygénation
cérébrale qu'il induirait
(Provine, 1987).
-
- Les pandiculations accompagnent
l'éveil; les bâillements
surviennent espacés durant la
journée, et accroissent leur
fréquence quand la pression du sommeil
augmente dans l'heure précédant
l'endormissement. Physiologiquement, un adulte
bâille de 5 à 15 fois par jour
(Baenninger, 1996; Walusinski, 2000). Comme il
existe des petits et des grands dormeurs, il
existe "des petits et des grands
bâilleurs". Les bâillements
apparaissent, en particulier, dans 4 situations
de la vie quotidienne: lire, voyager en
transport en commun, conduire, attendre (Greco,
1993).
-
- Les modifications d'état de vigilance
sont corrélées à des
variations de tonus musculaire antigravitaire.
Le système proprioceptif
(sensibilité profonde), par exemple des
trapèzes et des masséters, envoie
des influx par les fibres afférentes de
la catégorie Ia, situées dans la
racine mésencéphalique du
trijumeau. Celles-ci forment avec les
motoneurones des mêmes muscles une liaison
monosynaptique, base du réflexe
massétérin. La pandiculation et le
bâillement, par la contraction musculaire
massive, stimulent les structures responsables
de l'activation corticale, par les projections
de ces voies proprioceptives sur la formation
réticulée ascendante du tronc
cérébral et le locus coeruleus
(deux structures impliquées dans les
mécanismes de l'éveil),
anatomiquement proches du noyau du trijumeau
(noyau moteur des muscles manducateurs). A
noter, en corollaire, que l'amplitude du
réflexe massétérin varie
parallèlement au niveau de l'éveil
(Aubin et Garma, 1988).
-
- Le système nerveux
végétatif est un ensemble de
systèmes neuronaux afférents et
efférents régulant
l'homéostasie (Cannon, 1933), c'est
à dire le maintien à leur valeur
d'équilibre physiologique
différentes constantes corporelles:
température, pression artérielle,
osmolarité, etc. L'alternance jour/nuit
engendrée par la rotation terrestre
détermine, par l'intermédiaire des
noyaux supra-chiasmatiques de l'hypothalamus, la
composante circadienne de l'éveil et du
sommeil alors que l'hypothalamus (VLPO, noyau
paraventriculaire), des noyaux
mésopontins, la formation
réticulée ascendante du tronc
cérébral (locus coeruleus) et le
raphé antérieur régulent la
composante homéostasique. Il existe une
tendance à l'augmentation progressive de
l'activité vagale (parasympathique) de la
veille au sommeil lent avec pic lors du sommeil
paradoxal tandis que l'activité
sympathique décroît
parallèlement. Il en résulte une
hypotonie musculaire périphérique
généralisée et tendance au
collapsus des voies respiratoires
supérieures, caractéristiques du
sommeil paradoxal, prépondérant en
fin de nuit (Saper, 2005). L'intense
activité corticale propre à cette
phase de sommeil permet la transition vers
l'éveil (Campbell, 1985). La
pandiculation, et donc le bâillement,
ouvrent au maximum le pharyngo-larynx et
activent la reprise musculaire tonique,
déclenchant une augmentation de la
fréquence cardiaque, de la pression
artérielle, du métabolisme
musculaire associée à un
dérouillage articulaire. On peut
qualifier la pandiculation de
dynamogènique, terme utilisé par
Jean Lhermitte (1931).
-
- Ontogenèse.
- Le bâillement apparaît chez le
foetus simultanément à la
déglutition, témoignant de la
fonctionnalité du tronc
cérébral alors que la
myélinisation corticale n'est pas encore
acquise. Une ontogenèse chronologiquement
commune, la proximité anatomique des
centres exécutifs, témoins de
l'ancienneté phylogènique,
expliquent la synchronie des mouvements des
maxillaires et du cou. L'échographie en
3D animée (4D) permet leur visualisation
dès 12 à 14 semaines de grossesse
(Walusinski, 2005). Dès la naissance, ce
couplage fonctionnel est indispensable à
la survie, comme le montre l'effleurement du
coin de la bouche du nouveau-né qui
détermine une rotation
homolatérale de la tête, et permet
ainsi d'attraper le mamelon pour téter,
enclenchant le réflexe de
succion-déglutition. Ces
mécanismes neuro-moteurs
intégrés sont très stables
et ne varient pas au cours de la vie depuis la
naissance jusqu'à un âge
avancé (Humphrey, 1968).
-
- La vie foetale a un rythme ultradien de 90
minutes. Le bâillement indique la
transition entre un état
d'hypoactivité corrélé au
premier type de sommeil, forme immature de
sommeil paradoxal, encore
prépondérant à la
naissance, et un état d'activité
motrice intense (Feng, 2006).
-
- La phylogenèse suggère que
l'immobilité des poïkilothermes a
évolué vers le sommeil paradoxal
(Nicolau, 2000). La reprise de la
motricité des reptiles, dépendante
de l'élévation de la
température corporelle, peut
s'accompagner d'un bâillement. Ainsi,
l'ancienneté phylogénique et la
précocité ontogénique
indiquent un lien évolutif étroit
entre motricité bucco-cervicale,
bâillement et sommeil paradoxal.
-
- Ethologie.
- L'éthologie précise que le
bâillement est associé à
deux autres processus homéostasiques
contrôlés par l'hypothalamus, la
satiété et la
sexualité.
- La
satiété.
- Chez les animaux élevés en
captivité (rats, macaques, lions), il a
été noté des
bâillements répétés
précédant le moment des repas
proposés à heures
régulières, évoquant en
cela un rythme circadien (Holmgren, 1991).
- Les herbivores bâillent moins souvent
que les carnivores. Le rendement
énergétique de l'herbe est faible,
contrairement à celui de la viande. Les
herbivores passent un temps prolongé
à leur alimentation, dorment moins que
les carnivores et bâillent peu. Une
corrélation existe entre le nombre de
bâillements, la durée de sommeil,
en particulier du sommeil paradoxal, et le
niveau calorifique de la ration alimentaire. On
peut, dès lors, postuler que plus un
animal subit une pression vitale forte de
prédateurs, comme les herbivores, moins
il dort, moins il bâille. Plus il
ingère une ration calorique
élevée en peu de temps, comme les
carnivores prédateurs, plus il jouit d'un
luxe de sommeil prolongé et plus il
bâille. Seule exception, les primates non
humains, frugivores, ont une durée de
sommeil paradoxal et un nombre de
bâillements comparables à ceux des
carnivores (Halder et Schenkel, 1972).
- Chez l'homme, il est classique de ressentir
l'effet du jeûne par un fringale avec
bâillements (de même pour
l'hypoglycémie). Les excès
alimentaires, en particulier associés
à une alcoolisation
génèrent somnolence et
bâillements
répétés.
-
- La
sexualité.
- Chez les Macaques, que nous prendrons pour
exemple, les bâillements apparaissent
aussi après des interactions sociales
diverses en relation avec notamment la
sexualité ou des conflits.
L'éthologie montre que les mâles
bâillent plus souvent que les femelles. La
fréquence des bâillements augmente
avec l'apparition des caractères sexuels
secondaires (descente testiculaire, croissance
des canines), maximale chez les mâles
adultes. Le bâillement est sous la
dépendance partielle des
androgènes; la castration de macaques
adultes mâles entraînent une
diminution marquée de la fréquence
des bâillements tandis que des injections
de dihydrotestostérone exogène
rétablit la fréquence des
bâillements. L'injection d'un
anti-androgène non stéroïdien
(hydroxyflutamide) bloque les effets
d'injections de testostérone,
reproduisant les effets de la castration
(Deputte, 1994). Ces bâillements,
restreints aux mâles adultes et aux
mâles dominants dans le groupe, ont
été qualifiés de «
bâillements d'émotivité
» par Deputte (1980), soulignant ainsi
qu'ils sont déclenchés par une
« tension psychique ». Ils pourraient
servir à renforcer la place
particulière que l'individu occupe au
sein du groupe.
-
- Physiologie.
- Toutes les données
présentées permettent
d'interpréter pandiculations et
bâillements comme une activité
motrice extériorisée, d'origine
sous-corticale, témoignant de processus
homéostasiques régulés par
l'hypothalamus.
- Le noyau paraventriculaire de l'hypothalamus
est un centre intégrateur entre le
système nerveux central et le
système nerveux végétatif
périphérique. Un groupe de ses
neurones ocytocinergiques qui projettent sur des
aires extra hypothalamiques (hippocampe, tronc
cérébral, moelle) contrôlent
bâillements et érections. Leur
stimulation par la dopamine et ses agonistes,
des neuropeptides, l'ocytocine elle-même,
ou une stimulation électrique
déclenche des bâillements alors que
le GABA et ses agonistes et les opioïdes
inhibent bâillements et érections.
L'activation des neurones ocytocinergiques
nécessite une production d'oxyde nitrique
dont dépend la libération de
l'ocytocine extra-hypothalamique sans que les
mécanismes intimes soient actuellement
élucidés. D'autres
molécules modulent les bâillements,
non dans leur expression mais dans leur
fréquence et leur rythme:
sérotonine, hormones sexuelles,
hypocrétine, peptides endogènes
(ACTH, MSH). L'ocytocine stimule une activation
cholinergique au niveau de l'hippocampe, de la
réticulé ascendante du tronc
cérébral (Argiolas, 1994;
Sato-Suzuki, 1998). Enfin,
l'acétylcholine déclenche les
bâillements via les récepteurs
muscariniques des muscles effecteurs (noyaux des
nerfs V, VII, IX, X, XI, XII, C1-C4).
-
- La pandiculation et le bâillement
provoquent, à l'acmé de l'ample
inspiration, une augmentation de la pression
intra-thoracique, suivie d'une dépression
rapide, lors de l'expiration qui la suit. Il en
résulte un blocage du retour veineux et
lymphatique suivi d'un flux
accéléré. Il n'existe
actuellement aucune donnée sur les
conséquences immunitaires du drainage
lymphatique du canal thoracique vers la veine
cave, ainsi favorisé (Nolman, 2006). Par
contre, au niveau cérébral, la
variation rapide et notable de la pression
veineuse, accélère la circulation
du liquide céphalo-rachidien. En
conséquence, l'augmentation de la
clairance de l'adénosine, facteur
réputé somnogène,
interviendrait dans l'effet stimulant de
l'éveil du bâillement (Arrigoni,
2006).
-
- L'ample inspiration du bâillement
ouvre les alvéoles en atélectasie
et participe à l'étalement du
surfactant, comme le fait un soupir. Le
bâillement dirigé est ainsi
utilisé dans les suites
opératoires en chirurgie thoracique,
notamment (Cahill, 1978).
-
- Bâillements et
pathologies.
- La pandiculation et le bâillement ont
pour caractéristiques d'être
à la fois un symptôme clinique, un
traitement et aussi la cause de troubles
pathologiques.
-
- Le bâillement
symptôme clinique.
- La raréfaction ou la disparition des
bâillements, rarement spontanément
rapportée parce que peu gênante,
témoigne d'un syndrome parkinsonnien
(Goren, 1998), d'une consommation excessive de
caféine ou d'un effet opioïde. Dans
ces derniers cas, le sevrage s'accompagne d'un
rebond de bâillements, multiples, en
salves (Phillips-Bute, 1997). Le syndrome
d'Ondine congénital ou acquis
(malformations atloïdo-axoïdiennes)
témoigne du défaut fonctionnel du
contrôle de la ventilation par le tronc
cérébral (Ochoa-Sepulveda, 2005).
L'absence de bâillements, qui s'y attache,
l'oppose aux déficits corticaux
secondaires à une crise convulsive ou
à un accident vasculaire
cérébral où des
bâillements répétés,
d'origine diencépahlique, semblent
stimuler le cortex inhibé ou
lésé (Muchnik, 2003; Yankovsky,
2006).
-
- L'excès de bâillements, c'est
à dire des séries de 10 à
25 bâillements successifs,
irrépressibles,
répétés plusieurs fois par
jour, est actuellement le plus souvent d'origine
iatrogène et secondaire à la prise
d'anti-dépressifs
sérotoninergiques (Sommet, 2005). La
migraine s'accompagne fréquemment de
nombreux bâillements rappelant ceux
inaugurant l'installation d'un malaise vagal
(Cronin, 1988; Jacome, 2001). Les processus
expansifs intracrâniens, notamment la
pathologie adénomateuse hypophysaire,
certaines formes d'épilepsie temporale
peuvent s'accompagner de bâillements
répétitifs. Dans le cas
célèbre de JM. Charcot, la
patiente bâillait 8 fois à la
minute, soit 480 fois à l'heure,
était aménorrhérique, avait
une amputation bintemporale de son champ visuel
mais fut considérée comme
hystérique. La galactorrhée n'est
pas mentionnée mais ce cas évoque
un adénome à prolactine (Charcot,
1889). Les accidents vasculaires
cérébraux responsables de troubles
de la vigilance s'accompagnent de
fréquents bâillements (Cattaneo,
2006). Certaines hémiplégies
s'accompagnent d'un mouvements du bras
paralysé vers la bouche lors du
bâillement, curieux
phénomène que nous avons
décrit sous le nom de parakinésie
brachiale oscitante (Walusinski, 2005).
-
- Le bâillement
thérapeutique.
- A l'acmé d'un bâillement,
l'ouverture de la trompe d'Eustache aère
la caisse du tympan. Associée à
l'occlusion palpébrale, la baisse de
l'audition ainsi provoquée, concourt
à une brève déconnexion
sensorielle de l'environnement du
bâilleur. Lors de pathologies
rhino-pharyngées ou de variations
altudinales (montagne, vols aériens), le
bâillement constitue un moyen
thérapeutique de
reperméabilisation tubaire assurant
l'amélioration des surdités de
transmission, inflammatoires ou fonctionnelles
(Laskiewicz, 1953; Winther, 2005).
-
- La musculature extrinsèque et
intrinsèque de larynx est
particulièrement sensible au stress
émotionnel. Son hypercontraction est le
dénominateur commun de toutes les formes
de dysphonies et aphonies fonctionnelles ou
psychogènes. Le bâillement est une
thérapeutique des surmenages vocaux. Elle
est efficace pour combattre
l'élévation excessive du larynx et
la constriction de la glotte qui les
caractérisent. Ces surmenages
correspondent à un effort
tonico-spastique des muscles du larynx qui
provoque son ascencion. Le bâillement
ouvre la glotte à son maxima et
repositionne le larynx au plus bas de sa course,
réduisant l'effort musculaire. Les
professeurs de chant enseignent à leurs
élèves des techniques de
bâillements provoqués, forme de
relaxation laryngée. C'est ainsi qu'il
n'est pas rare de voir des chanteurs
d'opéra bâiller avant d'entrer en
scène (Xu, 1991; Boone, 1993).
-
- La laryngectomie totale a de multiples
effets secondaires, en particulier du fait de la
solution de continuité entre les voies
aériennes basses et hautes. Non seulement
la voix est perdue, mais aussi d'autres
fonctions physiologiques sont perturbées.
La perte de l'odorat s'ajoute aux
désagréments des modifications des
fonctions ventilatoires et à leurs
retentissements physiques et psychiques.
L'olfaction est tantôt un
phénomène passif survenant au
cours de la respiration, tantôt un
phénomène actif, inspiratoire et
volontaire. La laryngectomie totale supprime
l'olfaction passive, et seule une
minorité de patients est encore capable
d'olfaction volontaire. F. Hilgers (2002) a
développé une technique,
basée sur la création d'une
dépression dans la cavité buccale,
générant un flux d'air au travers
du nez. Les patients sont entraînés
à réaliser une imitation de
bâillement maximum mais en gardant les
lèvres strictement closes, tout en
aspirant leurs joues, abaissant leur langue et
le plancher de la bouche. La dépression
ainsi induite dans la cavité buccale
favorise une aspiration d'air par le nez
apportant les effluves parfumées.
-
- La théorie végétative
de James-Lange (1884-1885) postule que les
afférences en retour des contractions
musculaires et la sensibilité
viscérale apportent au système
nerveux central (striatum, amygdale, insula) les
perceptions corporelles, appelées
l'intéroception, permettant
l'élaboration consciente du schéma
corporel. La pandiculation et le
bâillement, par la puissante contraction
musculaire qu'ils représentent, la
déconnexion de l'environnement, et le
bien-être qui les suit, participent ainsi
de l'intéroception. Les techniques de
relaxation et de yoga utilisent implicitement
ces données, en déclenchant en
groupe, par échokinésie, des
bâillements induits, afin de perfectionner
le contrôle du tonus musculaire corporel
et une acutisation des perceptions
proprioceptives (Russell, 1891, Bourgne,
2006).
-
- Le bâillement inducteur
de pathologies.
- Le bâillement peut être le
déclencheur, la gâchette, d'une
crise de dystonie glossopharyngée, d'une
névralgie vasculaire de la face de
Charlin-Sluder (Jacome, 2001). Le syndrome de
Marin Amat ou syndrome de Marcus Gunn inverse,
consiste en une fermeture syncinétique
des paupières d'un oeil lors de
l'ouverture de la bouche notamment du
bâillement (Oh, 2003).
-
- Le bâillement peut déclencher
des crampes douloureuses des muscles
génohyoïdiens (André-Thomas,
1949).
- Par ordre de fréquence
décroissante, les causes de luxation
mandibulaire sont: le bâillement, le rire,
les vomissements, les soins dentaires. La
luxation est antérieure, le condyle
mandibulaire dépasse en avant le condyle
temporal et s'élève dans la fosse
temporale d'où il ne peut ressortir en
raison de la tonicité des muscles
masticateurs. Il s'agit le plus souvent d'une
luxation bilatérale (Ugboko, 2005). Une
prompte réduction peut
généralement être
réalisée sans anesthésie.
La récidive est fréquente.
Plusieurs publications rapportent des luxations
mandibulaires compliquant des excès de
bâillements irrépressibles sous
antidépressifs sérotoninergiques
(Tesfaye, 1990; Pae, 2003) ou lors d'induction
anesthésique (Avidan, 2002).
-
- Les fractures d'une longue apophyse
styloïde sont rares. Des douleurs de la
gorge ou du cou, une mobilité
réduite du cou, un enrouement, une masse
latérale du cou sont autant de signes
pouvant faire évoquer cette pathologie.
Elles sont secondaires à un
bâillement ou à un choc direct,
comme au cours des accidents de la circulation
(McGinnis, 1981; McCorkell, 1985). Il faut en
rapprocher le syndrome d'Eagle associant des
cervicalgies latérales,
déclenchées par les
bâillements, la mastication et à la
pression du processus styloïde dans la
fosse amygdalienne. La radiographie du cou
montre des calcifications des ligaments
stylo-hyoïdiens (découverte parfois
fortuite) qui deviennent symptomatiques avec ou
sans fracture (Mupparapu, 2005).
-
- Conclusion
- Bâillements et pandiculations sont des
comportements d'origine diencéphalique,
extériorisant un processus
d'homéostasie des systèmes
d'éveil (Walusinski, 2006). Sollicitant
puissamment les articulations
temporomandibulaires, ils conduisent vers le
stomatologue, notamment, les porteurs de
syndrome algo-dysfonctionnels de l'appareil
manducateur, les luxations de
mâchoire.
-
- Références
- Abrahams
VC, Richmond FJ. Specialization of
sensorimotor organization in the neck muscle
system. Prog Brain Res. 1988;76:125-35.
-
- Abrahams
VC, Kori AA, Loeb GE, Richmond FJ, Rose PK,
Keirstead SA. Facial input to neck
motoneurons: trigemino-cervical reflexes in the
conscious and anaesthetised cat. Exp Brain Res.
1993;97(1):23-30.
-
- André-Thomas,
de Ajuriaguerra J. Etude sémiologique
du tonus musculaire. Flammarion Ed, Paris 1949.
844p.
-
- Argiolas
A, Melis MR. The neuropharmacology of
yawning. Eur J Pharmacol. 1998;343(1):1-16.
-
- Arrigoni
E, Chamberlin NL, Saper CB, McCarley RW.
Adenosine inhibits basal forebrain cholinergic
and noncholinergic neurons in vitro.
Neuroscience. 2006;140(2):403-413.
-
- Aubin HJ,
Garma L. Le bâillement. Psych
Psycho-biol. 1988;3:275-286.
-
- Avidan
A. Dislocation of the temporomandibular
joint due to forceful yawning during induction
with propofol. J Clin Anesth.
2002;14(2):159-160.
-
- Baenninger
R, Binkley S, Baenninger M. Field
observations of yawning and activity in humans.
Physiol Behav. 1996;59(3):421-4225.
-
- Barbizet
J. Le bâillement. Concours Med.
1958;80(5):537-538.
-
- Boone DR,
McFarlane SC. A critical view of the
yawn-sigh as a voice therapy technique. J Voice.
1993;7(1):7580..
-
- Bourgne
N. Bâiller c'est quoi? Quel sens en
yoga? Mémoire d'enseignant de yoga, Ecole
Alsacienne de Yoga. Strasbourg. 2006.
-
- Cahill CA.
Yawn maneuver to prevent atelectasis. AORN J.
1978;27(5):1000-1004.
-
- Campbell
SS. Spontaneous termination of ad libitum
sleep episodes with special reference to REM
sleep. Electroencephalogr Clin Neurophysiol.
1985;60(3):237242.
-
- Cannon
WB (1871-1945). The wisdom of the body.
Norton and Company, New York. 1932, 333p.
-
- Cattaneo L,
Cucurachi L, Chierici E, Pavesi G.
Pathological yawning as a presenting symptom of
brain stem ischaemia in two patients. J Neurol
Neurosurg Psychiatry. 2006;77(1):98-100.
-
- Charcot
JM.(1825-1893) Bâillement
hystérique. Leçons du mardi
à La Salpêtrière. Lecrosnier
& Babé Ed. Paris. 1889;2:1-11.
-
- Cronin TG.
Yawning: an early manifestation of vasovagal
reflex. AJR Am J Roentgenol.
1988;150(1):209.
-
- Deputte
BL. Ethological study of yawning in primates
quantitative analysis and study of causation in
two species of Old World Monkeys (Cercocebus
albigena and Macaca fascicularis). Ethology.
1994;98:221-222.
-
- Deputte BL,
Johnson J, Hempel M, Scheffler G. Behavioral
effects of an antiandrogen in adult male rhesus
macaques (Macaca mulatta). Horm Behav.
1994;28(2):155-164.
-
- Duchenne
de Boulogne GB (1806-1875). Mécanisme
de la physionomie humaine. Jules Renouard
Libraire. Paris 1862. 194p.
-
- Feng
P. The developmental regulation of
wake/sleep system. In Neuroendocrine correlates
of sleep/wakefulness. Cardinali DR and
Pandi-Perumal SR Ed. Springer Ed. New York.
2006. 3-18.
-
- Forte MC.
Le bâillement. Thèse Doctorat en
médecine, n°117. Bordeaux. 1982.
89p.
-
- Goren JL,
Friedman JH. Yawning as an aura for an
L-dopa-induced "on" in Parkinson's disease.
Neurology. 1998;50(3):823.
-
- Greco
M, Baenninger R, Govern J. On the context of
yawning: when; where; and why ? Psych Rec.
1993;43:175-183
-
- Halder U,
Schenkel R. Das Reich-Gähnen bei
Rindern (Bovinae). Zeitschrift für
Säugetierkunde. 1972;37:232-245.
-
- Heloe
B, Heloe LA. Frequency and distribution of
myofascial pain-dysfunction syndrome in a
population of 25-year-olds. Community Dent Oral
Epidemiol. 1979;7(6):357-360.
-
- Hilgers
FJ, Jansen HA, Van As CJ, Polak MF, Muller MJ,
Van Dam FS. Long-term results of olfaction
rehabilitation using the nasal
airflow-inducing("polite yawning") maneuver
after total laryngectomy. Arch Otolaryngol Head
Neck Surg. 2002;128(6):648-54.
-
- Holmgren
B, Budelli R, Urba-Holmgren R, Eguibar JR,
Holmgren M, Baz-Tellez G, Anias J. Food
anticipatory yawning rhythm in the rat. Acta
Neurobiol Exp (Wars). 1991;51(3-4):97-105.
-
- Humphrey
T. The development of mouth opening and related
reflexes involving the oral area of human
fetuses. Ala J Med Sci. 1968;5(2):126-157.
-
- Jacome DE.
Compulsive yawning as migraine premonitory
symptom. Cephalalgia. 2001;21(5):623-625.
-
- Jacome DE.
Primary yawning headache. Cephalalgia.
2001;21(6): 697-699.
-
- James
W (1842-1910). What is an emotion? Mind.
1884;9:188-205.
-
- Lange
KG (1834-1900). Om Sindsbevægelser et
psyko-fysiologisk Studie. Lund Ed.
Kjøbenhavn. Denmark. 1885, 91p.
-
- Laskiewicz
A. The role of the Eustachius tube and the
tympanal muscles in yawning. Rev Laryngol Otol
Rhinol. 1953;74(5-6):309-316.
-
- Lhermitte
J. Le sommeil. Armand Colin Ed, Paris 1931.
212p.
-
- Marder
E, Rehm KJ. Development of central pattern
generating circuits. Curr Opin Neurobiol.
2005;15(1):86-93.
-
- McCorkell
SJ. Fractures of the styloid process and
stylohyoid ligament: an uncommon injury J
Trauma. 1985;25(10):1010-1012.
-
- McGinnis
JM Jr. Fracture of an ossified stylohyoid
bone. Arch Otolaryngol.1981;107(7):460.
-
- Muchnik
S, Finkielman S, Semeniuk G, de Aguirre MI.
Yawning and temporal lobe epilepsy Medicina (B
Aires). 2003;63(2):137-139.
-
- Mupparapu
M, Robinson MD. The mineralized and
elongated styloid process: a review of current
diagnostic criteria and evaluation strategies.
Gen Dent. 2005;53(1):54-59.
-
- Nicolau
MC, Akaarir M, Gamundi A, Gonzalez J, Rial
RV. Why we sleep: the evolutionary pathway
to the mammalian sleep. Prog Neurobiol.
2000;62(4):379-406.
-
- Nolman B.
The Lymphatic pump. Yawnology. 2006.
http://ob1.fcpages.com/
-
- Ochoa-Sepulveda
JJ, Ochoa-Amor JJ. Ondine's curse during
pregnancy. J Neurol Neurosurg Psychiatry.
2005;76(2):294.
-
- Oh JY,
Kim JE, Park Kd, Choi KG. A case of familial
inverse Marcus Gunn phenomenon. J Neurol
Neurosurg Psychiatry. 2003;74(2):278.
-
- Pae CU, Kim
JJ, Lee CU, Lee SJ, Lee C, Paik IH. Injured
temporomandibular joint associated with
fluoxetine-monotherapy-induced repeated yawning.
Gen Hosp Psychiatry. 2003;25(3):217-218.
-
- Phillips-Bute BG,
Lane JD. Caffeine withdrawal symptoms
following brief caffeine deprivation. Physiol
Behav. 1997;63(1):35-39.
-
- Provine RR,
Tate BC, Geldmacher LL. Yawning: no effect
of 3-5% CO2, 100% O2, and exercise. Behav Neural
Biol. 1987;48(3):382-393.
-
- Russell
H. Yawning. The Delsarte Series n°1.
John W. Lovell Company Ed, New York. 1891.
175p.
-
- Saper
CB, Cano G, Scammell TE. Homeostatic,
circadian, and emotional regulation of sleep. J
Comp Neurol. 2005;493(1):92-98.
-
- Sato-Suzuki I,
Kita I, Oguri M, Arita H. Stereotyped
yawning responses induced by electrical and
chemical stimulation of paraventricular nucleus
of the rat. J Neurophysiol.
1998;80(5):2765-2775.
-
- Sommet A .
Drug-induced yawning: a review of the French
pharmacovigilance database. Fundamental Clin
Pharmacol. 2005;19(2):227.
-
- Tesfaye Y,
Lal S. Hazard of yawning.
CMAJ.1990;142(1):15.
-
- Ugboko VI,
Oginni FO, Ajike SO, Olasoji HO, Adebayo ET.
A survey of temporomandibular joint dislocation:
aetiology, demographics, risk factors and
management in 96 Nigerian cases. Int J Oral
Maxillofac Surg. 2005;34(5):499-502.
-
- Walusinski
O. Pourquoi bâillons-nous? Rev Prat
Méd Gén.
2000;14(487):259-263.
-
- Walusinski
O, Kurjak A, Andonotopo W, Azumendi G. Fetal
yawning assessed by 3D and 4D sonography. The
Ultrasound Rev Obs Gyncecol.
2005;5(3):210-217.
-
- Walusinski O.
Yawning: Unsuspected avenue for a better
understanding of arousal and interoception. Med
Hypotheses. 2006;67(1):6-14.
-
- Winther
B, Gwaltney JM Jr, Phillips CD, Hendley JO.
Radiopaque contrast dye in nasopharynx reaches
the middle ear during swallowing and/or yawning.
Acta Otolaryngol. 2005;125(6):625-628.
-
- Xu
JH, Ikeda Y, Komiyama S. Bio-feedback and
the yawning breath pattern in voice therapy: a
clinical trial. Auris Nasus Larynx.
1991;18(1):67-77.
-
- Yankovsky
A, Andermann F, Dubeau F. Post-ictal
forceful yawning in a patient with nondominant
hemisphere epilepsy. Epileptic Disord.
2006;8(1):65-69.
-
- Zafar
H, Nordh E, Eriksson PO. Temporal
coordination between mandibular and head-neck
movements during jaw opening-closing tasks in
man. Arch Oral Biol. 2000;45(8):675-682.
-
- Somnolter©
développé par Nomics (http://www.nomics.be)
- WSL-2 Liège Science Park
- rue des Chasseurs Ardennais, 4
- B-4031 Angleur (Belgium)
-
|
