Department of Zoology, North
Carolina State University, Raleigh
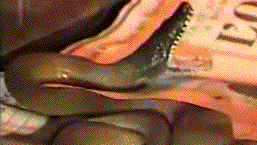
African clawed frogs fed to American water
snakes induced yawning and gaping which
slowed ingestion and facilitated the frogs'
escape without inducing flavor aversion. The
peptide and/or indolealkylamine contents of the
frog's poison glands caused the effect because
frogs with purged glands did not induce these
behaviors and rarely escaped. Poison gland
mucus, applied orally, elicited similar oral
movements. The frog's clear lubricating mucus
was inactive. As several compounds in the poison
glands have known neuroleptic properties, the
oral behaviors may be induced by neural
mechanisms reported to govern
neuroleptic-induced orofacial dyskinesia in
schizophrenics.
The skin secretions of many amphibians
contain peptides that are identical to, or close
analogues of, peptides found in the vertebrate
brain and gut. Despite studies on peptides that
form this "brain-gut-skin triangle", little is
known of their role within amphibian skin. As
poison glands of the African clawed frog
(Xenopus !aevis) contain only peptides and
indolealkylamines, we explored the potential
antipredatory role of these compounds by feeding
Xenopus to American northern water snakes
(Nerodia sipedon). Here we report that such
feedings cause dyskinetic yawning and gaping
movements in the snake that frequently permitted
a frog's escape. The dyskinesia is predictable
given the known actions of each skin compound
and their coincidence with neural mechanisms
believed to underly drug- and peptide- induced
oral dyskinesias in mammals including man.
RESULTS
Only toxic mucus consistently induced
yawning and gaping. Fixed yawns
and gapes (lasting >4.0 sec) were common in
toxic mucus trials. Snakes yawned and gaped
usually after having climbed the tank walls.
Some fixed yawns and gapes occurred under
water suggesting that yawning and gaping
need not be associated with breathing. We
conclude that toxic mucus induced involuntary
yawning and gaping and permitted frogs to escape
because few oral behaviors appeared in snakes
given other mucus treatments.
DISCUSSION
Xenopus skin mucus contains the
indolealkylamines, serotonin (5-HT) and
bufotenidine (BF), and the following peptides:
cholecystokinin octapeptide (CCK-8); caerulein
(CRL), a close structural and functional
analogue of CCK-8; r thyrotropin-releasing
hormone (TRH); and xenopsin (XN), an analogue of
neurotensin. Curiously, the action of each
compound is compatible with hypotheses governing
the tardive dyskinesias (TD) seen in
schizophrenics treated chronically with
dopaminergic blockers (the neuroleptics).
TD is an extrapyramidal dysfunction in which
involuntary yawning, chewing and tongue
movements occur. However, as dopamine inhibits
release of tuberoinfundibular peptide hormones,
the neuroleptics also elevate plasma levels of
prolactin and oxytocin, alphamelanocyte
stimulating hormone and adrenocorticotropin,
which have all induced yawning in lab
animals. As CCK-8 and CRL and 5-HT have
neuroleptic properties, and XN may possess the
neuroleptic properties of neurotensin, we
suspect that these Xenopus skin compounds mimic
neuroleptics and create a neurochemistry for
oral dyskinesia in snakes that promotes the
escape of frogs.
Further, 5-HT and bufotenidine and TRH
elevate serum prolactin levels and oxytocin, the
most potent known inducer of yawning, is
dramatically elevated in plasma of rats
administered CCK.
Our observation that oral dyskinesias occur
within 30 seconds of oral contact with mucus
suggests that toxins are absorbed orally and/or
react with oral receptors. The snake's
vomeronasal organ (VNO), a contact receptor that
mediates detection of prey and the rate of
tongue flicking, is probably not the chief
target for skin toxins because two snakes given
VNO nerve transections (but unconfirmed
histologically) exhibited all presurgical
dyskinesias.
This new behavioral model of orofacial
dyskinesia appears biomedically promising given
that the known compounds in Xenopus skin occur
naturally in the mammalian brain or occur as
close analogues of neurochemicals. Whether skin
agents act in combinations or alone is the focus
of our future studies. We are now assessing the
behavioral responses of Lycodonomorphus rufulus,
a natural snake predator of Xenopus in South
Africa (J. Visser, personal communication).
Typically, sympatric predator and prey species
coevolve adaptations to each other's defenses.
Thus, if African water snakes prove unresponsive
to Xenopus' skin mucus, and if this defense is
neurologic, such as an unusual profile of
neurotransmitters typically affected by
neuroleptics, rapid advances in drug development
can be made by comparing the brain chemistry of
Nerodia with that of Lycoodonomorphus.


 video
video