In a study of human yawning Robert
Provine (1986) remarked that yawning is a
prominent stereotyped action pattern and
releasing stimulus which "does not deserve its
current status as a minor behavioural
curiosity". Indeed, "yawning may have the
dubious distinction of being the least
understood, common, humnan behaviour". In
contrast, a large body of experimental data has
been collected during the past 30 years on
drug-induced yawning in animals
(particularly rodents).
In this chapter we consider the neural basis of
drug-induced yawning in rodents and discuss the
relevance of this pharmacological phenomenon to
"spontaneous" yawning in animals and man.
We propose that yawning rmay be controlled
by a complex interaction of catecholaminergic,
serotonergic, and peptidergic neuronal
mechanisms. A model is put forward to explain
how yawning may be caused largely by peptidergic
and cholinergic excitation and dopaminergic
inhibition. Furthermore, we present evidence
which suggests that, in animals and man, yawning
may be a marker of recovery from acute stress
and that these responses may be closely
associated with an inhibition of brain dopamine
metabolism. [...]
Interaction of brain dopaminergic,
cholinergic, and peptidergic neurones in the
mediation of yawning: an hypothesis and a
model
It is clear from the preceding pages that
yawning behaviour is influenced by a number of
interacting; neurotransmitter systems. Major
influences are exerted by brain dopaminergic,
cholinergic, and peptidergic neurones and in
this section we propose a hypothesis which
explains how these various neurotransmitter
mechanisms may interact to control yawning and
associated behaviours.
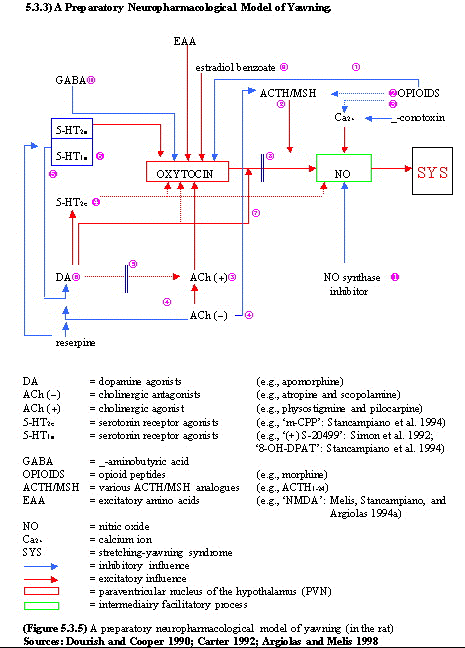
There is strong evidence for the involvement
of dopaminergic inhibition and cholinergic
excitation in yawning. Thus, yawning induced by
dopaminergic drugs is probably caused by
activation of dopamine autoreceptors (or
inhibitory postsynaptic dopamine receptors)
which reduces dopamine synthesis and release. In
contrast, yawning induced by cholinergic agents
appears to be due to increased release of
acetylcholine and stimulation of postsynaptic
muscarinic receptors.
In cross-blocking studies, it bas been shown
that dopamine agonist induced yawning; is
attenuated or abolished by treatment with
muscarinic receptor antagonists (Yamada
and Furukawa 1980, 1981; Holmgren
and Urba-Holmgren 1980). In contrast,
dopamine antagonists potentiate yawning induced
by physostigmine (Yamada and Furukawa 1980;
Holmgren and Urba-Holmgren 1980). Therefore,
these authors have proposed that yawning is
produced by the release of cholinergic neurones
from tonic dopaminergic inhibition. This
disinhibition may be caused by activation of
dopamine autoreceptors at presynaptic neuronal
sites induced by low doses of dopamine agonists.
Thus, the same functional effect (ie. increased
yawning) is produced by stimulation of
postsynaptic cholinergic receptors or
presynaptic (inhibitory) dopamine receptors.
Blockade of postsynaptic dopamine receptors by a
dopamine antagonist would also activate
cholinergic neurones and this probably accounts
for the potentiation of physostigmine-induced
yawning by neuroleptics (Yamada and Furukawa
1980).
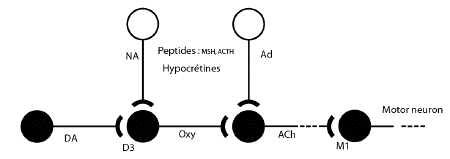
It seems that the striaturm may be the
central locus for this dopaminergic-cholinergic
neuronal interface. The striatum in the rat
receives innervation from approximately 3500
dopaminergic neurones located in the zona
compacta of the substantia nigra (Andén
et al. 1964, 1966). The terminals of these
dopamine neurones make synaptic connections with
striatal interneurones (which represent the
majority of striatal neurones) and neurones
which innervate the substantia nigra and globus
pallidus. Acetylcholine is a major striatal
transmitter and most of it is located in
interneurones (Hassler 1978). The dopaminergic
nigrostriatal neurones make synaptic contact
with these cholinergic neurones (Hattori et al.
1976) and inhibit their firing (Roth and Bunney
1976; Trabucchi et al. 1975). In contrast, there
is no apparent dopaminergic-cholinergic link in
other major dopamine terminal regions such as
nucleus accumbens and olfactory tubercles
(Ladinsky et al. 1975).
In yawning experiments, it has been shown
that the striatum is very sensitive to dopamine
agonist treatments and that 6-OHDA lesions of
the striatum. or substantia nigra abolish
yawning induced by a small dose of apomorphine
(Dourish et al. 1985; Dourish and Hutson 1986;
Stoessl
et al. 1987). Furthermore, yawning induced by
intrastriatal injection of piribedil is
abolished by blockade of either dopamine
autoreceptors (with low-dose haloperidol) or
postsynaptic muscarinic receptors (with
scopolamine) (Dourish et al. 1985). At this
point, our yawning model comprises striatal
cholinergic excitation and dopaminergic
inhibition. We noted earlier that the peptide
hormones ACTH, a-MSH, beta-LPH and oxytocin are
potent yawning inducers. Therefore, the question
arises as to how peptidergic mechanisms interact
with neuronal dopamine and acetylcholine to
control yawning.
There is evidence that ACTH and alpha-MSH
injection can activate cholinergic neurones
(Torda and Wolff 1952; Marx 1975). Accordingly,
yawning induced by ACTH and alpha-MSH is
paralleled by a twofold elevation of
acetycholine turnover in hippocampus (Wood et
al. 1978). This is consistent With evidence that
peptide-induced yawning is suppressed by
cholinergic antagonists and neuroleptics
(Ferrari et al. 1963; Yamada and Furukawa 1981).
These data suggest that yawning is associated
with cholinergic and peptidergic excitation and
dopaminergic inhibition.
Indeed it is known that alpha-M S
H-producing cells in the pituitary are under the
inhibitory control of dopaminergic neurones
originating from the arcuate nucleus of the
hypothalamus (Tilders and Smelik 1977).
Therefore, it is possible that inhibition of
dopamine release (caused by low-dose dopamine
agonist treatment) may indirectly result in
release of newly synthesized peptides (ACTH,
alpha-M S H, betaLPH, oxytoxin) from the
pituitary or from peptidergic neurones
(Serra et al.
1983a).
The importance of the pituitary in mediating
yawning is illustrated by the observation that
hypophysectomy prevents yawning induced by
apomorphine (Serra et al. 1983a). Similarly,
apomorphine-induced yawning is prevented by
treatment with the protein synthesis inhibitor
cycloheximide (Serra et al. 1983b).
The observation of Wood et al. (1978) that
ACTH and alpha-MSH specifically increased
acetylcholine turnover in the hippocampus
indicates that this brain region may be of
importance in the control of yawning. This idea
is supported by evidence from lesion studies in
which it has been demonstrated that partial
ablation of the hippocampus potentiates
ACTH-induced yawning whereas total
hippocampectomy abolishes the response (Colbern
et al. 1977). This study also implicated the
amygdala and the mammillary bodies in the
control of yawning since lesions in these areas
modified the response to ACTH.
Interestingly, lesion studies have also
enabled the differentiation of the yawning and
sexual arousal components of the ACTH-induced
syndrome. Thus, pre-optic lesions, destroying
structures which take up labelled testosterone,
abolished penile grooming and erection but did
not affect yawning (Bertolini et al. 1975).
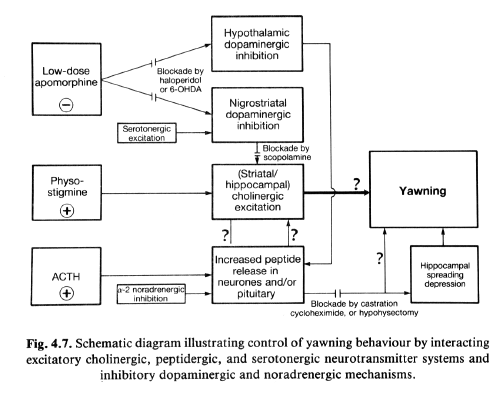
The model we propose to explain the neural
control of yawning is illustrated in Fig. 4.7.
It is clear that there are cholinergic,
peptidergic, serotonergic (all excitatory),
dopaminergic, and noradrenergic (both
inhibitory) inputs to the system. At this point
it is unclear whether the final step in the
pathway is peptidergic or cholinergic. However,
it is noteworthy that all of these influences
may precede a mechanism illustrated on the
bottom right portion of Fig. 4.7. Cortical
spreading depression was shown to produce
yawning and sexual arousal by Huston (1971). In
a subsequent study, Jakobartl and Huston (1977)
observed that intracranial injection of ACTH
produced spreading depression and that the
hippocampus was more sensitive to the peptide
than the cortex. Thus, it is possible that
yawning and related behaviour elicited by ACTH
could be secondary to hippocampal spreading
depression.
Conclusions
In this chapter we have described how
drug-induced yawning is mediated by the
interaction of various brain neurotransmitter
systems. Dopaminergic, peptidergic, and
cholinergic neurones appear to be primarily
responsible for the control of yawning. At the
pharmacological level yawning and sexual arousal
appears to be useful as a model for identifying
drugs with agonist activity at inhibitory
dopamine receptors (Gower et al. 1984).
Similarly, chewing mouth movements have been
proposed as a useful index of agonist action at
central muscarinic receptors (Salamone et al.
1986).
In behavioural terms, the evidence suggests
that in most cases pharmacologically-induced
yawning bears a close resemblance to
spontaneous, physiological yawning. Thus, the
posture of rats yawning in response to
physostigmine or apomorphine is very similar to
that of spontaneous physiological yawning in
rats (Ushijima
et al. 1985). Furthermore, apomorphine induced
yawns in rats occur in clusters (Szechtman 1984;
Cooper, de Mars, and Dourish, unpublished
results) which is consistent with anecdotal
reports that yawning in humans occurs; in bures
(Barbizet
1958).
The only comprehensive study to date on
physiological yawning in animals was carried out
by Anias
et al. (1984) who produced a "high yawning
frequency" line of Sprague-Dawley rats through
selective breeding. They found a clear circadian
pattern in spontaneous yawning with the highest
frequency being evident during the last hour of
the light period. Interestingly, this coincides
with the time of the lowest daily dopamine
turnover rate (Cahill and Ehret 1981) which
suggests some form of dopaminergic control of
spontaneous yawning.
Spontaneous, physiological yawning is a
behaviour categorized by ethologists as a
"stereotyped action pattern" (see Provine
1986 and references therein). In humans, yawns
can be released by observing yawns, thinking
about yawning, or even reading about yawning
(Provine 1986). There have been a number of
speculations concerning the function of yawning.
One proposal is that yawning is useful for
"stretching the face". By causing contraction of
the facial muscles, yawning forces blood through
cerebral vessels to the brain which may have an
alerting effect (Heusner
1946; Barbizet
1958). Similarly, it has been suggested that
yawning may increase blood oxygen levels during
the deep air inspiration which accompanies the
response (Bartlett
et al. 1971). However, a study by Provine (1986)
has cast doubt on the respiratory hypothesis
since there was no correlation between yawn
duration and interyawn interval (ie. infrequent
yawners did not compensate by producing yawns of
longer duration).
Yawning is also of clinical interest since
it has been reported to be associated with a
number of disorders including epilepsy, epidemic
encephalitis and Huntington's chorea (Heusner
1946), hysteria and brainstem lesions
(Barbizet 1958) and adrenoleucodystrophy (which
interestingly is accompanied by high blood ACTH
levels; Kataoka et al. 1980). Yawning is also
reported to be associated with opiate withdrawal
in man (Himmelsbach 1939). In contrast, it has
been reported that apomorphine-induced yawning
in rats is reduced by the opiate antagonist
naloxone (Szechtman 1984). However, it appears
that the effects of naloxone on yawning may not
be opiate-receptor-mediated (Berendsen
and Gower 1986).
It seems likely that yawning may have an
important social function both in apes (Hadidian
1980) and in humans (Barbizet 1958). In man,
yawning is often regarded as an expression of
indifference and/or boredom although social
etiquette demands that the yawn is hidden by
putting one's hand over one's mouth.
We believe that yawning and stretching may
signal the termination of stressful experience
or of sustained concentration. Experimental
evidence is lacking, but there are anecdotal
observations which suggest that yawning and
stretching may be behavioural features of
recovery from at least certain forms of stress.
For example, one of us (SJQ sat amongst a large
class of students in Northern Ireland who were
being addressed by a visiting rescarch worker.
He wanted them to answer direct questions about
the impact of the "Troubles'" (ie. the period
from 1969 to the present day during which there
has been widespread violence) on their personal
lives, and on those of their families and
friends. The effect of his talk on the behaviour
of the students was startling. Under normal
circumstances, like students anywhere, his
audience would have shown periodic fidgeting,
whispering, looking-about, coughing, and so on.
On this occasion, however, the entire class sat
motionless, and expressionless, when it became
clear that they were being asked about
widespread fears and anxieties, and about
injuries and deaths which may have befalien
members of their families, their friends, and
neighbours. There was a palpable feeling of
tension throughout the lecture room.
As the speaker came to an end of his talk
and signalled this by some closing comments, the
behaviour of the class changed remarkably. They
relaxed their body postures, they turned to
classmates and looked at each other, they
smiled, and most strikingly, there was a
widespread outbreak of yawning and stretching.
These changes were closely synchronized
throughout the class. It was difficult to
discount the impression that the yawning and
stretching occurred as part of a more complex
change in the students' behaviour, which was
initiated by the end of a distressing
experience. Formal observations of the behaviour
of people, alone or in large groups, during and
following the imposition of stress would be
extremely interesting. We suggest that the
occurrence of yawning and stretching, in people,
may form part of a range of behavioural
responses indicative of recovery from stressful
events. The animal data which we discuss imply
that in people, too, yawning and stretching may
follow from neurochemical changes in the brain,
which include an inhibition of central
dopaminergic activity. In rats it is clear that
spontaneous yawning; can be significantly
altered by environmental manipulation. In
animals and man changes in brain dopamine
metabolism and yawning frequency may be closely
associated with recovery from acute stress.
During the past decade experiments on
drug-induced yawning; in animals have
facilitated that construction of a model of the
neuronal circuity which subserves yawning.
Furthermore, yawning in animals has proved to be
a useful pharmacological tool for studying
neurotransmitter receptors and receptor
subtypes. The challenge remains to discover the
physiological trigger for yawning and to fully
understand the behavioural and social
significance of the response.
-Banks
RJ, Mozley L, Dourish CT. The angiotensin
converting enzyme inhibitors captopril and
enalapril inhibit apomorphine-induced oral
stereotypy in the rat. Neuroscience.
1994;58(4):799-805
-Stoessl
AJ, Dourish CT, Iversen SD
Apomorphine-induced yawning in rats is abolished
by bilateral 6-hydroxydopamine lesions of the
substantia nigra Psychopharmacol; 1987; 93;
336-342
-Stoessl
AJ Effects of ageing on the behavioural
responses to dopamine agonists: decreased
yawning and locomotion, but increased stereotypy
Brain Research 1989; 495; 20-30
-Stoessl
AJ, Dourish CT, Iversen SD The NK-3
tachykinin agonist senktide elicits yawning and
chewing mouth movements following subcutaneous
administration in the rat. Evidence for
cholinergic mediation. Psychopharmacology
(Berl). 1988; 95; 4; 502-506
-Cooper SJ;
Dourish CT Neural basis of drug induced
yawning in neurobiology of stereotyped behaviour
Oxford ed 1990
-Cooper
SJ, Rusk IN, Barber DJ. Yawning Induced by
the Selective Dopamine D2 Agonist N-0437 is
Blocked by the Selective Doparnine Autoreceptor
Antagonist (+)-UH 232. Physiol Behav.
1989;45:1263-1266
-Dourish
CT, SJ Cooper Yawning elicited by sytemic
and intrastriatal injection of piribedil and
apomorphine in the rat Pyschopharmacology 1985;
86; 175-181
-Dourish
CT, PH Hutson Bilateral lesions of the
striatum induced with 6-hydroxydopamine abolish
apomorphine-induced yawning in rats
Neuropharmacology 1985; 24; 11; 1051-1055
-Collins
G, JM Witkin et al Dopamine agonist-induced
yawning in rats: a dopamine d3 receptor mediated
behavior J Pharmacol Exp Ther 2005