Two different neurotransmitters,
acéehylcholine (ACh) and dopamine (DA)
have been recently postulated to neurohumoral
mechanisms, underlying the act of yawning. One
group of investigators working on physostigmine-
and pilocarpine-induced yawning in infant rats,
has stressed the role of central muscarinic
cholinergic synapses, because the yawning effect
induced by both cholinomimetic drugs is blocked
by scopolamine. On the other hand, Mogilnicka
and Klimek have described that low doses of
systemically injected DA agonists in adult rats
produce recurrent episodes of yawning, responses
which are completely inhibited by the DA
antagonist spiperone. Basically similar
observations have been later reported by Di
Chiara et al.
As physostigmine-induced. yawning, both in
infant and in 45-dayold rats, is strongly
potentiated by previous administration of Lu
10-171 (citalopram), a potent and selective
serotonin uptake blocking drug, a suggestion has
also been advanced that serotonin (5-HT) may
exert a modulating effect on yawning behavior
serotonergic-modulation.html.
Some questions immediately arise when these
results are considered. Are the ontogenetic
curves of yawning induced by cholinergic or
dopaminergic drugs similar or different? May
cholinergic and dopaminergic pathways somehow
interact in the induction of yawning? Does
serotonin also modulate
dopaminergically-elicited yawning?
The experiments presented in this paper try
to give an answer to these questions in the hope
that a unified hypothesis regarding the
neurohumoral basis of yawning may eventually
emerge. This seems to be a relevant problem,
since it has been shown that several other
conditions or drugs are able to induce yawning:
intraventricular infusions or intracerebral
injections of adrenocorticotrophic hormone
(ACTH) or fragments of ACTH, spreading
depression of the neocortex and hippocampus, and
administration of naloxone. [...]
Discussion : After Mogilnicka
and Klimeik's demonstration that yawning may
be elicited in adult rats by the administration
of dopaminergie drugs, our previous suggestion
that yawning might be a
cholinergic response needed re-examination.
Cowan had also, pointed out that other factors,
aparat from cholinergic mechanisms, could induce
yawning. He recalled that i.m. administration of
dimethyltryptamine causes yawning in rhesus
monkeys. DPI, (3,4
dihydroxyphenylamino)-2-imidazoline, which has
been described to have specific and potent
agonistic activity on DA inhibitory receptors,
also elicits yawning in infant rats.
In the present experiments, using
apornorphine in the same low dose range as
Mogilnicka and Klimek, we have confirmed their
and Di Chiara results, showing that this drug
definitely elicits yawning in adult male albino
rats. We have also traced the ontogenetic
course of this effect. Apomorphine does not
induce yawning in 7-day-old rats, does it only
slightly before the eleventh day, but quite
distinctly by the 15th day, when the effect
praetically reaches the same level observable
until the end of the first month. A 50% higher
average effect is obtained in adult rats
(3-month-old). Increase with age of oher
behavioral effects of DA agonists has recently
been reported. The ontogenetic evolution of
apomorphine-elicited yawning thus follows an
absolutely different curve than the one which
illustrates the same behavior evoked by
physostigmine or pilocarpine.
Cholinomimetically-induced yawning is
highest during the first seven postnatal days,
falling thereafter to a level which is not
significantly different from spontaneous yawning
on the 21st day. The different maturation course
of cholinergic- and dopaminergic-induced yawning
responses naturally suggest different underlying
mechanisms. Nevertheless, the fact that both
cholinomimetic- and apomorphineelicited yawning
are blocked by low doses of scopolamine, points
to the decisive importance of some muscarinic
cholinergic synaptic links in the
neuroanatomical circuits subserving yawning. How
could one envisage the coupling of dopaminergic
and cholinergic synapses in order that yawning
might be elicitable both by dopamino- or
cholinomimetic agents and subject to block by
scopolamine whatever the eliciting drug? One
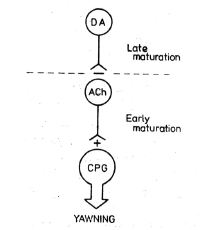
possible explanation is offered by the tentative
and simple model presented in Fig. 6, in which
DA and ACh synapses appear organized "in
series", the latter exerting an excitatory
influence on the central pattern generator of
yawning behavior, and thus being able to trigger
the response. The early maturation of yawning,
both spontaneous and cholinornimetically-induced
justifies the suggestion that the central
pattern generator "yawning center" and its
cholinergic trigger may have a quite caudal
location in the brainstem. This is in keeping
with the observation of yawning in anencephalic
humans, with the opinion of the early reviewers
of yawning behavior and the most recent
description of yawning in a completely
tetraplegic patient suffering from a
transecting glioma of the pons.
Figure 6 also suggests that the cholinergic
yawn-triggering neurons are under the control of
later maturing inhibitory dopaminergic neurons.
In rats, their gradual restraining influence on
yawning behavior may be illustrated by the
declining tendency of cholinomimetic yawning
from around the 7th postnatal day onwards, which
corresponds roughly to the reciprocal evolution
of apomorphine-elicited yawning. In agreement
with previous authors we interpret apomorphine
induced yawning as the result of activation of
low threshold presynaptic inhibitory DA
receptors, "autoreceptors" of Carlsson, by low
doses of apomorphine, thus liberating the
cholinergic neurons triggering yawning from a
tonic dopaminergic restraining influence. Higher
doses of apomorphine or other DA agonists, by
directly stimulating high threshold postsynaptic
DA inhibitory receptors on the cholinergic
neurons, would exert an inhibitory effect on
yawning. The dose-effect of apomorphine-induced
yawning demonstrates the very narrow dose range
of this effect, which begins to decline already
with 0.20 mg/kg. This seems to be the reason why
authors studying stereotyped activity with
higher doses of apomorphine failed to observe
its yawning-eliciting action. If this
interpretation is correct it could be expected
that if excessive apomorphine is injected (for
an optimal yawn-eliciting effect), and the
concentration of the drug falls, as a result of
metabolic degradation, late yawning activity
might appear. We have confirmed this expectation
in a group of 5 young adult rats injected with
apomorphine (1 mg/kg). In that case significant
yawning appeared only in the second half of a 60
min observation period while in rats injected
with 0.05 mg/kg yawning began already 5 min
after injection and had practically disappeared
after half an hour (unpublished
observations).
The "crossed blocking" experiment, in which
physostigmine induced yawning is increased after
blocking DA receptors with spiroperidol, while
the same behavior elicited by apomorphine is
decreased, provides further evidence in support
of the idea that an important dopaminergic tonic
inhibitory influence on yawning is present
around the 15th postnatal day in the rat. The
potentiating effect of spiroperidol on
physostigmine-induced yawning is not significant
in rats 7 to 11 days old, in concordance with
the relative immaturity of the dopaminergic
influences on yawning, as illustrated by the
ontogenetic course of apomorphine-elicited
yawning.
An important serotonergic facilitatory effect
on physostigmine-induced yawning has been
previously described. If DA and ACh synapses
influencing yawning were organized as suggested
in Fig. 6, it would be reasonable to expect that
serotonin uptake inhîbitors should have a
similar effect on apomorphine-elicited yawning
as they have on that behavior evoked by
physostigmine. Our experiments testing the
action of citalopram and fluoxetin on yawning
induced by apomorphine failed to demonstrate
such effect. This is contradictory to the
demonstration that a significant depression of
apomorphine-elicited yawning is obtainable with
metergoline and, therefore, difficult to
explain. Thus, the idea that serotonergic
pathways may exert a general tonic facilitatory
or modulatory influence on yawning, although
appealing, should perhaps require further
substantiation.
The exploration of the effect of drugs
modifying dopaminergic and cholinergic synaptic
transmission upon yawning induced by other
methods may help in testing the more general
validity of the tentative model of organization
of the synaptic links regulating and triggering
yawning behavior presented in this work (Fig.
6)