Barbizet
was perhaps right in suggesting that the
commonplace, ubiquitous and apparently
non-significant of the act of yawning, might
have contributed to the relative lack of
attention given by physiologits to this
behavioural phenomenon. The physiological
signifiance of yawning and the nervous
mechanisms triggering and coordinating its
various components remain unclear. Frequent
yawning has been mentioned as a symptom in some
diseases of the central nervous system,
particularly in cases of frontal tumours and
encephalitis. A stretching and yawning syndrome
has been observed in cats, dogs and monkeys
after intracisternal or intraventricular
injections of adrenocorticotrophic and
melanotrophic hormones, or synthetic
polypeptides with ACTH and/or MSH activities,
but the relation of these observations to the
basic mechanism of yawning has not been stutied.
During experiments designed to explore
cholinergic potentiation of d-amphetamine
induced head-shaking in infant rats, we have
made some observations that indicate the
involvement of central cholinergic response in
yawning.
Infant rats, from 6h after birth up to 14 d
old, were ijected intraperitoneally (ip) with
physiostigmine salicylate (0,1 mg of the free
base per kg body weight, dissolved in 0,9%NaCl
to a total volume of 0,01 ml per g body weight).
The rats began to yawn about 5 mn after
injection. In the following 15 mn the number of
yawns reached an average of 10-12 during first
10 d of life. Some animals yawned twice per
minutes, and others only a couple of times in 15
mn. In rats aged under 10d physiostigmine
induced yawning was practically universal.
Between days 10 and 14 of life the frequency of
yawning fell rather steeply, to a levelof only 2
or 3 yawns per 15 mn, with some animals showing
no yawning reponse. At the 14th day yawning
occurence had decreased to 69% of the
population. Each yawn was preced by salivation,
licking of the forepaws and cleaning movements
of the snout, or "gomming" or chewing movements,
and sometimes by a sudden stretching of the
forelimbs. The yawn itself was a slow wide
opening of the mouth, with marked protrusion of
the toungue, and lasted about 3-4 s.
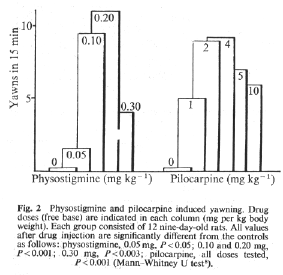
A dose effect graphe of physiostigmine
induced yawning in 9d old rats is show fig 2.
That this particular action of the
anticholinesterasic drug is centrally mediated
is strongly suggested by control experiments
with neostigmine methylsulphate, which passes
the blood-brain barrier with difficulty. When
injected i.p., in a dose of 0.2 mg per kg,
neostigmine induced only 1.6 yawns per 15 min
(12 animals). Although this effect is
significantly different from that observed in
rats injected with saline (P<0.01) it was
only of the order of one-sixth of the effect
obtained with physostigmine. With higher doses
of neostigmine no yawning was observed.
Pilocarpine hydrochloride, a cholinomimetic drug
that passes readily into the brain, induced
yawning up to the same level as
physostigmine.
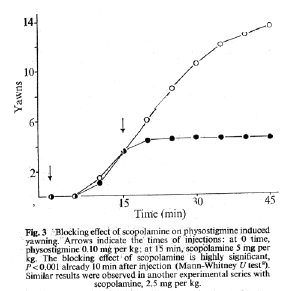
Cholinomimetically induced yawning seems to
be due to the stimulation of central
'muscarinic' receptors ' because it is rapidly
and completely blocked by scopolamine
hydobromide, at doses of 2.5 to 5 mg per kg
(Fig3). That 'nicotinic' cholinergic receptors
are not participating was demonstrated with
nicotine administered by i.p. injection. Doses
of nicotine (0.1 mg/kg) which induced clear
central effects on head-shaking (unpublished
observations) did not induce yawning.
We observed physostigmine induced yawning at
a more or less stable level of 2-3 yawns per 15
mn in rats from 14 to 90 d of age, but
practically only in males. In a pool of 62
animals (31 females and 31 males, 14 to 5.4 d
old) from the same litters, males performed an
average of 3.1 yawns in 15mn after physostigmine
injection, whereas females, performed 0.5 yawns
in the same period. The difference is highly
significant, (P<0.002) by the Mann-Whitney U
test. It may be recalled that Hall, in his
interesting description of the bebaviour of
monkey towards mirror-images, in which "yawning"
or "gasping" responses were very frequent, also
observed that adult females "rarely, if ever
yawned".
Our results suggest that in the rat thre is a
central cholinergic mechanism, with muscarinic
receptors, underlying the act of yawning. The
mechanism matures at a very early age,
suggesting that it is localised at the lower
levels of the brainstem. Later development of
higher placed cholinergic mechanisms may
determine that other effects evoked by
cholinomimetic agents injected systemically
might be hindering the expression of yawning in
olders rats. Intense and continous scratching,
or tremor or rigidity, as produced respectively
by pilocarpîne and physostigmine in rats
aged over 10d, or some other central or
peripheral actions affecting the level of
vigilance, might be partially counteracting the
yawn-inducing effect.
Nature vol 271 12 january 1978
Cholinergic link in
yawning A. COWAN Department of
Pharmacology,Temple University School of
Medicine, Philadelphia, Pennsylvania 19140
Holmgren et al, recently focused attention on
the basic mechanism behind the act of yawning.
They reported that physostigmine and pilocarpine
induce yawning in young male rats and
hypothesised that acentral cholinergic link may
be involved in the reflex. Our results support
surch proposal.
Yawning is a characteristic sign of
with-drawal from morphine in man and monkeys.
When naloxone (0.5 mg per kg bodyweight), but
not physiological saline, was injected
subcutaneously (sc) into three ex-addict baboons
(two male and one female, 4.4 - 5.2 kg), 98 d
after abrupt with-drawal of morphine, a low
incidence of yawning (2-4 episodes) occurred
within 15 min; on this occasion, other signs of
long-term withdrawal were absent. Seven days
later, the same baboons were again challenged
with naloxone, 20 mn after physostigmine (0.05
mg per kg s.c.). Although this dose of
physostigmine per se did not elicit yawning,
with each animal there was a threefold increase
in the incidence of yawning.
Although a cholinergic link may indeed be
involved in yawning, it should be recognised
that other factors are also important. Thus,
dimethyltryptamine causes yawning after
intramuscular administration t0 rhesus monkeys
yet there is no evidence that this hallucinogen
has marked effects on the cholinergic system in
this species.
Urba-Holmgren and Holmgren reply
In Cowan's experiments 'with exmorphine
addict baboons, although physostigmine 0.05 mg
pet kg s.c. per se did net evoke yawning, this
dose strongly potentiated naloxone-induced
yawning. Perhaps a higher dose of physostigrnine
(0. 10 mg per kg) would have elicited yawning
directly even in monkeys, as it does in rats and
in infant guinea pigs, kittens and dog pups
(unpublished observations). In infant rabbits we
have had te use a still higher dose (0.15 - 0.20
mg per kg).
We certainly agree with Cowan that other
factors are also important in yawning. The
stretching and yawning syndrome induced by ACTH
and MSH is a well-known example. In recent
experiments with (3,4
dihydroxyphenylamino)-2-imidazoline (DPI,
Böhringer) we have observed that this drug,
which according to Cools et al has a specific
and potent agonistic activity at dopamine
inhibitory receptors, in doses of 5 mg per kg
intraperitoneally, elicits moderate, but
statistically significant yawning in infant rats
(from 9 to 15 d in age). It would berash at this
stage, to hypothesise that drugs shown te elicit
the yawning act necessarily through a final
common path including a cholinergic link.
Eguibar JR
et Moyaho A Inhibition of grooming by
pilocarpine differs in high-and low yawning
sublines of Sprague-Dawley rats. Pharmacoology
Biochemistry and Bebavior 1997; 58: 2
317-322
Eguibar
JR et al Behavioral differences between
selectively bred rats: D1 versus D2 receptors in
yawning and grooming Pharmacology, Biochemistry
and Behavior 2003; 74; 827Ð832
Moyaho A et
al Induced grooming transitions and open
field behaviour differ in high and low-yavning
sublines of Sprague-Dawley rats. Anim Behav
1995; 50 ; 61-72
Urba-Holmgren R,
Trucios N, Holmgren B, Eguibar JR, Gavito A,
Cruz G, Santos A Genotypic dependency of
spontaneous yawning frequency in the rat Behav
Brain Res 1990 Oct 30;40(1):29-35
Urba-Holmgren
R, Santos A, Holmgren B, Eguibar JR Two
inbred rat sublines that differ in spontaneous
yawning behavior also differ in their responses
to cholinergic and dopaminergic drugs Behav
Brain Res 1993 Sep 30;56(2):155-9
Urba-Holmgren
R, Gonzalez RM, Holmgren B Is yawning a
cholinergic response? Nature 1977 May 19 267
(5608): 261-2 et commentaires Cholinergic link
in yawning A Cowan Nature 12/01/78 271
p187-188