Pendant les
bâillements (Bouyssou
1985), on note un ralentissement des
vitesses du flux
jugulaire plus qu'une
accélération, avec
élévation de la pression veineuse
intracranienne. Cela a-t-il un effet sur la
résorption du liquide
céphalorachidien par les plexus
choroïdes ? La conséquence en est
actuellement inconnue (action
sur la clairance de l'adénosine et
des prostaglandines hypnogènes PGD2 ?),
mais la physiologie de la circulation du LCR est
actuellement remise
en cause. (Lire: Schroth
G and Klose U Cerebrospinal fluid flow;
Physiology of respiration-related
pulsations)
Le bâillement comme toute inspiration
profonde augmente le retour veineux au coeur
droit, avec augmentation de la pression dans
l'artère pulmonaire (Woodbury).
FH Lepp (1982) a
bâti une théorie sur le rôle
du bâillement comme pompe veineuse
auxilliaire, par compression du plexus veineux
ptérigoïdien. En
2015, Ertekin et al. ont montré la
smilitude életrophysiologique entre
bâillement et déglutition: les
mêmes centres bulbo-pontiques sont
activés et chaque bâillement est
suivi d'une déglution.
Le massif facial et le cerveau
s'individualisent à partir d'une
structure embryonnaire commune : l'ectoblaste.
Le pôle céphalique comporte
une
segmentation originelle embryologique (voir
photos de foetus de 44 et
54 jours) encéphalo-faciale et
encéphalo-cervicale avec une
correspondance topographique stricte : les
structures naso-frontales et
prémaxillaires sont liées au
cerveau antérieur; les structures
maxillo-mandibulaires et cervicales
antérieures sont unies au tronc
cérébral et à ses nerfs. Au
début du troisième mois, l'embryon
devient un fœtus grâce à
l'apparition des premières
séquences motrices orales et
pharyngées sous la dépendance du
développement neurologique du tronc
cérébral: développement de
l'activité de succion-déglutition,
bâillement. Succion et bâillement
ont donc la même origine
embryologique, soulignant l'importance du
tronc cérébral dans le
développement neuro-physiologique de
l'activité oro-pharyngée
coordonnée avec les régulations
respiratoire, cardiaque et digestive de
même localisation neuro-anatomique. Le
bâillement et la succion sont
détectables chez le foetus humain,
à l'échographie, dès
la 12° semaine de grossesse, stade
où le tronc cérébral est
individualisé et l'hypophyse devient
fonctionnelle, alors que l'extension du
néocortex temporal et frontal se
complète jusqu à 22 à 24
semaines.(voir bâillement
foetal)
Pendant les quelques heures que peuvent
vivre des anencéphales,
il a été noté qu'ils
bâillaient et s'étiraient. Des
patients atteints de locked-in
syndrome, bien que paralysés,
bâillent encore (voir
expériences du chat
décortiqué). Le syndrome
operculaire
(syndrome de
Foix-Chavany-Marie) réalise une
dissociation automatico-volontaire, avec
paralysie faciale centrale, linguale et
pharyngolaryngée mais la bâillement
reste possible. L'étirement par
l'examinateur d'un
membre en hypertonie pyramidale d'un
hémiplégique peut parfois
déclencher un bâillement. Ou,
à l'inverse, le bâillement peut
entrainer une élévation du membre
supérieur paralysé (voir Thèse
Dr Quoirin) La
Parakinésie Brachiale
Oscitante.
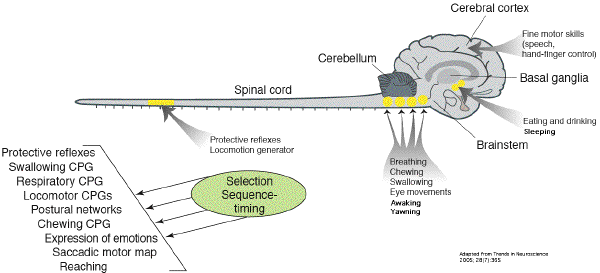
La
régulation du sommeil et de
l'éveil est sous la dépendance
d'une quinzaine de circuits différents et
redondants, situés principalement dans le
pont (adrénergique), dans le
pédoncule (dopaminergique), dans
l'hypothalamus (histaminergique), dans la
région basifrontale de Meynert
(cholinergique). Il
existe des réseaux permissifs
contrôlant l'éveil qui doivent
être inhibés pour que le sommeil
apparaisse. Le déclenchement du
bâillement pourrait intervenir "en effet
latéral" de ces réseaux.
(voir schéma des connexions de
l'hypothalamus.) L'influx issu des fuseaux
neuromusculaires massétérins
sensibles à l'étirement semble
avoir également un rôle dans le
déclenchement du processus (voir
nouvelles conceptions). Le bâillement
est souvent perçu comme une jouissance,
un bref bien-être, ressemblant aux
satisfactions des tiqueurs.Cette perception est
médiée par les voies
spécifiques de l'intéroception.
Les
progrès dans la compréhension des
systèmes neurobiologiques de l'alternance
éveil/ sommeil permettent aussi de
proposer une nouvelle théorie, où
le bâillement serait le trait
comportemental révèlant la
transition d'un état à l'autre :
théorie
du l'interrupteur flip - flop
(switch).
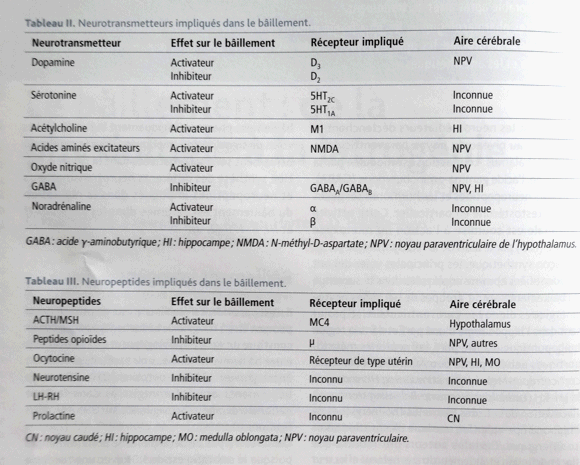
La participation du système
dopaminergique peut-être mise en
évidence par l'administration de
faibles doses
d'apomorphine,
agoniste
mixte des récepteurs D1-D2-D3
des synapses
dopaminergiques qui induit des
bâillements. De fortes doses les font
disparaître au profit de
stéréotypies motrices chez
l'animal ou de dyskinésies
essentiellement du faciès chez l'homme.
Ces bâillements induits par l'apomorphine
( ou le pirebedil)
sont antagonisés
par les neuroleptiques typiques et atypiques,
mais pas par la dompéridone, agent
bloqueur des récepteurs dopaminergiques
périphériques car ne diffusant pas
à travers la barrière
hématoencéphalique. Un
agoniste D1 pure ne déclenche pas de
bâillements, quelque soit la dose. Ceux-ci
paraissaient donc induits par la stimulation des
récepteurs
D2 à seuil d'excitation
bas (récepteurs
de forte affinité), liés
aux
récepteurs D1 dont la
stimulation est indispensable pour obtenir
un bâillement.
L'affinement
des technologies permet, en 2007, une remise en
cause de cette conception. L'équipe
de G.
Collins
(Ann Arbor) considère le bâillement
comme un comportement très
spécifique de l'activation
des
récepteurs
D3,
très probablement au niveau du
noyau
paraventriculaire de
l'hypothalamus.
Le nombre de bâillements induits par des
agonistes dopaminergiques dessine une courbe en
cloche inversée (inverted U-shape).
L'induction des bâillements dépend
des récepteurs D3 et à fortes
doses une inhibition compétitive avec les
récepteurs D2 engendrerait une
réduction du nombre des
bâillements. (The results of these
experiments provide convergent evidence that
dopamine D2/D3 agonist-induced yawning
is
a D3 agonist mediated
behaviors,
with subsequent inhibition of yawning being
driven by competing D2 agonist activity).
(FICY
2010).
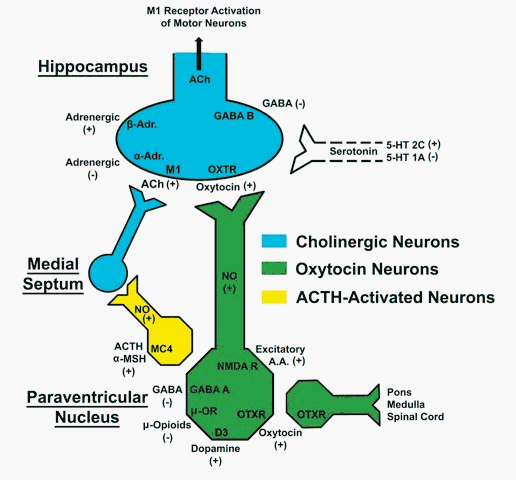
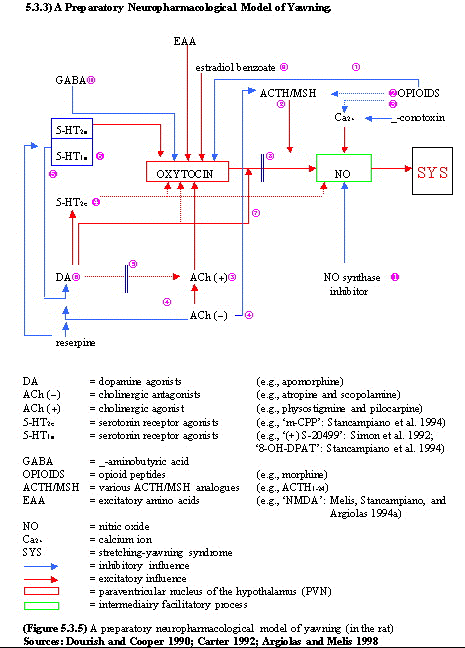
Systèmes des peptides
endogènes: L'hypophysectomie
fait disparaitre les bâillements.
L'ACTH, a-MSH
(Melanotonine
stimulating hormon) (qui dérivent
comme les endorphines de la
pro-opiomélanocortine), la LH-RH,
soit trois petides hypophysaires injectés
chez le rat en intrathécal provoquent
bâillements et érection.
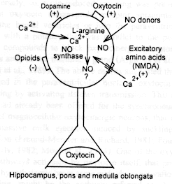
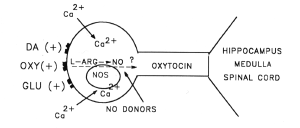
L'ocytocine injecté dans le noyau
paraventriculaire (sa destruction empêche
tout bâillement) de l'hypothalamus
déclenche des bâillements, alors
qu'un inhibiteur de l'ocytocine (vasotocine) les
inhibe.
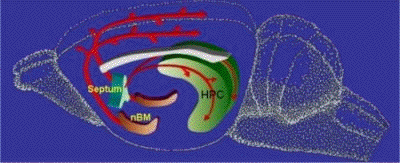
Le rôle de l'hypothalamus et de
l'hypophyse
dépend d'un réseau hypothalamique
ocytocinergique,
au niveau paraventriculaire
(Sato-Suzuki) qui
reçoit des influences activatrices
dogaminergiques, et des influences
inhibitrices opioïdes. Ce réseau
projette sur l'hippocampe d'une part et la
région bulbopontique d'autre part (celle
probablement qui est exécutive du
bâillement). On retrouve au niveau de ce
noyau
hypothalamique un parallèle entre
activation ou non du bâillement et la
présence ou non d'oxyde
nitrique synthétase. Enfin notons que
les antagonistes du glutamate provoquent
étirements et bâillements:
d'où l'idée d'essayer le
baclofène
à action GABAergique comme
traitement
des salves de bâillements. (essai jamais
réalisé à ce jour).
La découverte
en 1998 de l'hypocretine,
neuro-médiateur
sécrété par l'hypophyse
latérale complète le rôle
important de l'hypophyse. Les données
concernant la prolactine
sont limitées mais paraissent
liées aux voies dopaminergiques. Enfin,
il semble exister une modulation alpha et beta
adrénergique; les
béta-bloquants capables de franchir
la barrière
hémato-encéphalique, augmentent la
fréquence des bâillements.
Bâillements
et benzodiazépine.
Les travaux de neurobiologie
de l'attachement (mère-enfant;
individu-groupe, individu-individu)
dévoile le rôle
prépondérant des cellules du noyau
praventriculaire sécrétant de
l'oxytocine : une piste pour comprendre le
bâillement
d'émotivité ?
L'influence sérotoninergique
est révèlée par diverses
expérimentations pharmacologiques. Le
mCPP
(1-3-chlorophenyl-piperazine) sélectif
des récepteurs 5-HT2c est un puissant
inducteur des bâillements aussi bien chez
l'homme que l'animal. Ceci explique,
probablement, l'effet iatrogène de salves
de bâillements chez des patients qui
reçoivent des agonistes
sérotoninergiques comme
antidépresseurs. Par contre la
stimulation des récepteurs 5-HT1A et
5-HT2 empêche l'apparition des
bâillements induits aussi bien par
l'apomorphine que par le mCPP ou les inhibiteurs
de l'oxyde
nitrique (NO,
substanceP).
Le noyau paraventriculaire de l'hypothalamus,
qui reçoit d'importantes projections
sérotoninergiques depuis le noyau du
raphé dorsal, serait le siège
d'une
modulation des neurones dopaminergiques et
ocytocinergiques
par ces différents types de
récepteurs sérotoninergiques
qu'ils portent. (voir
schéma)
L'histamine
intervient dans le maintien de l'éveil
par activation corticale depis les noyaux
tubéro-mammillaires.
L'histamine déclenche les
bâillements ce qui est un argument
indirect pour évoquer un effet stimulant
des bâillements.
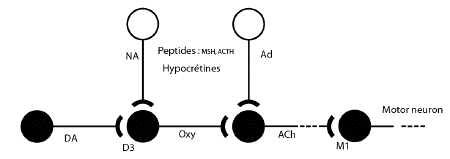
En résumé : il semble exister
une voie mésolimbique dopaminergique
(noyau A10 - septum) et une voie cholinergique
(septum
- hippocampe).
A noter que les
variations circadiennes cholinergiques et
dopaminergiques concordent avec celles du
bâillement chez le rat.
Chez les mammifères sauf l'Homme,
bâillement et érection sont
associés. Les mécanismes
physiologique ne sont pas
élucidés. Le
bâillement est
testostérone-dépendant
dans de nombreuses espèces
animales mais pas chez l'homme. A noter le
rôle des
aires préoptiques latétales
dans le déclenchement de
l'érection associée au sommeil
paradoxal.
L'hypocrétine,
découverte en 1998, régule
à partir du noyau paraventriculaire
hypothalamique la satiété et
l'éveil. La place de l'hypocrétine
dans la neurophysiologie du bâillement
commence à apparaitre
expérimentalement chez le rat :
l'injection d'hypocrétine 1 dans le
noyau parventriculaire de l'hypothalamus
déclenche des bâillements en
stimulant la vigilance. L'hypocrétine 2
stimule la vigilance sans déclencher de
bâillements : voir Sato-Suzuki
I, H Arita Cortical arousal induced by
microinjection of orexins into the
paraventricular nucleus of the rat .
L'observation montre que les carnivores
bâillent plus que les herbivores, comme
les variations du niveau de leur vigilance le
laissent prédire (Baenninger). En effet,
le rythme de vie des carnivores est
caractérisé par d'importantes
variations du niveau de vigilance, depuis la
tranquilité somnolente, à la
marche, la course après une proie, la
chasse, le dépeçage. Au contraire,
la vie des herbivores est relativement monotone,
et ils passent la plupart de leur temps à
brouter; la difficulté à fabriquer
des protéines à partir de l'herbe
obligent ces mammifères à passer
beaucoup plus de temps de leur éveil
à en ingérer.Seuls les primates
non humains, essentiellement frugivores, contre
disent cette règle, la fréquence
de leurs bâillements étant proche
de celle des carnivores. On peut supposer ici
un rôle régulateur de
l'hypocrétine sur le sommeil et
l'homéostasie énergétique
et donc sur la fréquence des
bâillements.
-Sanna
F, Succu S, Melis MR, Argiolas A. Dopamine
agonist-induced penile erection and yawning:
Differential role of D(2)-like receptor subtypes
and correlation with nitric oxide production in
the paraventricular nucleus of the hypothalamus
of male rats. Behav Brain Res. 2012
Les muscles du bâillement
Muscles de la face ou muscles de la mimique
(38)
Muscles superficiels (25)
Muscle occipito-frontal (ou
épicrânien, ventre frontal et
occipital), impair, innervé par le nerf
facial
Muscles auriculaires (muscles
antérieur, supérieur et
postérieur)
Muscle orbiculaire de l'œil (parties
orbitaire, palpébrale et lacrymale)
Muscle procérus (muscle pyramidal),
impair
Muscle élévateur (commun) de
la lèvre supérieure et de l'aile
du nez (muscle naso-labial ou grand
sus-maxillo-labial)
Muscle petit zygomatique
Muscle grand zygomatique
Muscle orbiculaire de la bouche, impair
Muscle risorius
Muscle abaisseur de l'angle de la
bouche
Muscle mentonnier
Muscles profonds (13)
Muscle corrugateur du sourcil,
innervé par le nerf facial
Muscle nasal (parties transverse et
alaire)
Muscle abaisseur du septum nasal,
impair
Muscle buccinateur
Muscle élévateur (propre) de
la lèvre supérieure (muscle moyen
sus-maxillo-labial)
Muscle abaisseur de la lèvre
inférieure
Muscle élévateur de l'angle de
la bouche (muscle canin)
Muscles de l'appareil manducateur ou muscles
manducateurs (8)
Muscle masséter (parties profonde et
superficielle), innervé par le nerf
massétérique
Muscle ptérygoïdien
médial, innervé par le nerf
mandibulaire
Muscle ptérygoïdien
latéral, innervé par le nerf
mandibulaire
Muscle temporal
Muscles supra-hyoïdiens (8)
Muscle stylo-hyoïdien, innervé
par le nerf facial
Muscle génio-hyoïdien
Muscle mylo-hyoïdien, innervé
par le nerf mandibulaire
Muscle digastrique (digastrique
génio-hyo-mastoïdien), ,
innervé par le nerf mandibulaire