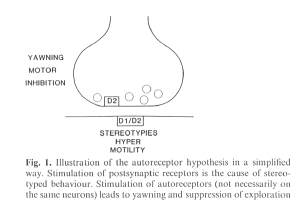
Discussion : we will firt discuss the
data underlying the autoreptor hypothesis and
then continue with the present results,
alternative interpretations and the clinical
perspective.
Data underlying the autoreceptor
hypothesis : The original finding, which led
to the formulation of the autoreceptor
hypothesis (as defined in the introduction), was
that APO inhibits tyrosine hydroxylation (dopa
accumulation in NSD 1015 - treated mice) in the
same doses as motor activity of mice is
suppressed. This result has been confirmed
beyond doubt by many investigators. An extension
of this evidence is provided by Brown, who
showed that inhibition of tyrosine hydroxylation
(dopa accumulation in GBL- and NSD 1015-treated
rats) by a wide range of DA agonists correlate
nicely with suppression of exploration in mice.
However, it is uncertain to what extent the NSD
method reflects changes in synaptic levels of
DA. Thus, Brown found that alphaMPT (250 mg/kg,
1 h) did not alter the behavioural effects of DA
agonists which is compatible with our findings.
Their result was interpreted as evidence that DA
agonists suppress exploration by a mechanism
independent of inhibition of synthesis. This
certainly agrees with our finding that doses of
alphaMPT (50-100 mg/kg) which reduce
extracellular DA more than a behaviourally
active dose of APO (0.05 nig/kg) have no effect
on behaviour. Thus, we conclude that inhibition
of synthesis is not the mechanism by which DA
agonists suppress exploration or induce yawning.
These findings demonstrate that the evidence
used to formulate the autoreceptor hypothesis in
the first place is indirect and relies upon the
assumption that changes in synthesis are
followed by similar changes in release.
Other results supporting the autoreceptor
hypothesis are that lesions of the striatum or
the substantia nigra by 6-OH DA or in the
substantia nigra by electrocoagulation abolish
yawning induced by APO given systemically.
However, Scheel-Krüger found that yawning,
induced by local injection of DA agonists into
the striatum, was not abolislied by 6-OHDA
lesions in the same region. Recently Melis
reported that APO injected locally into the
hypothalamus elicit yawning in very low doses (5
ng). Hence, it cannot be excluded that this
discrepancy is due to non-specific effects of
the larger lesions. It should be noted that
neither lesion was reported to induce yawning,
as may be expected since both autoreceptor
stimulation and 6-OHDA lesions should result in
a reduced DA transmission.
It has also been suggested that the
antagonistic effect of reserpine (4 h) on
APO-induced
yawning supports the autoreceptor hypothesis.
Surprisingly, the finding that reserpine (18 h)
potentiates yawning induced by APO was also
regarded as evidence in favour of the
autoreceptor hypothesis. Serra et al. found that
reserpine (18 h) alone induces yawning in an
alphaMPT- and sulpiride-sensitive manner, which
was regarded as evidence against the
autoreceptor hypothesis. Hence, the effects of
reserpine on APO-induced yawning are not
unimpeachable evidence in favour of the
hypothesis.
Data questioning the autoreceptor
hypothesis : Behavioural evidence that low
doses of DA agonists have effects similar to
postsynaptic DA receptor stimulation is provided
by the fact that suppression of exploration and
induction of stereotypies are not two extremes
on the same scale. The use of a method where
several beliavioural variables can be recorded
simultaneously is a prerequisite for this
observation. It is well known that recording of
some variables can give seriously misleading
results when comparisons between different
devices are attempted. By combining the
multivariate registration with multivariate
statistical analysis one ensures that, given the
limitations of the recording devices, changes in
behavioural patterns are detected. Thus, we
conclude that an "activity-inactivity" scale
does not account for the dose-response to DA
agonists.
Compounds that are D2 receptor agonists or
mixed D1/D2 agonists suppress exploration in
various models. The D1 partial agonist SKF 38393
has also been found to suppress exploration,
although this finding is controversial. We found
that SKF 38393-induced suppression of
exploration was not antagonised by sulpiride.
Thus, we conclude that suppression of
exploration is D2 receptor mediated.
In addition, we have found that suppression
of exploration can only be antagonised by
D2-receptor blocking agents. The selective D1
antagonists SCH 23390 has no effect on
APO-induced suppression of exploration. In
contrast, the selectiveD2 receptor antagonist
sulpiride completely antagonises this response
in a wide dose range. The antagonistic effects
by other DA receptor blockers, e.g. haloperidol
and cis-flupenthixol are more controversial.
Some authors report antagonistic properties
of haloperidol while others find no effect.
Similarily, cis-flupenthixol has beenreported to
be eiher antagonistic or to have no effect.
Taken together, many neuroleptics can, in a
small dose range, antagonize suppression of
exploration induced by DA agonists in some
models in some species. Two D2 antagonists stand
out as unique. Remoxipride has no antagonistic
effect on APO-induced suppression of exploration
and sulpiride is the only substance that is an
effective antagonist over a wide dose range
(Stàhle and Ungerstedt 1986b, 1987b). In
addition, the antagonisrn by sulpiride is
surmontable. We have also found that
extracellular levels of sulpiride in the dose
range corresponding to the in vivo Kd of 5-10
mg/kg does not exceed 10 nM. These data suggest
that, e.g. haloperidol- and remoxipride-induced
suppression of exploration is mediated by a
different population of D2 receptors than
sulpiride. Thus, it is concluded that DA
agonist-induced suppression of exploration is
mediated by a subclass of D2-receptor.
An interesting observation is that
haloperidol and sulpiride both cause an increase
in extracellular DA, DOPAC and HVA and levels of
DOPAC and HVA are increased by remoxipride. From
this it may be concluded that the receptors
mediating increases in extracellular DA, DOPAC
and HVA are pharmacologically distinct from
those mediating suppression of exploration.
These observations, together with our early
finding that there was discrepancy in the
time-course of the action of APO on
extracellular DA and suppression of exploration
prompted us to test the autoreceptor hypothesis.
In particular, the four predictions listed in
the introduction were investigated by combined
microdialysis and behavioural studies. Our
conclusions based on the results obtained from
these tests are:
1. Reduction of extracellular DA to a given
level by drugs with different pharmacological
mode of action does not result in similar
behavioural changes. Inhibition of tyrosine
hydroxylase by MPT reduced extracellular levels
of DA in microdialysis experiments. The effects
of MPT (50-100 mg/kg) were of at least the same
magnitude as the effects of APO (0.05 mg/kg).
However, these doses of MPT had no effect on
exploration and did not induce yawning.
Reserpine (2 mg/kg) reduced DA levels to
approximately the same extent as APO (0.05
mg/kg). However, 3 h after injection of
reserpine there were no behavioural effects of
the drug while after 4 h there was a strong
suppression of exploration. Similar comparisons
hold for pergolide and EMD 23448.
2. An increase in extracellular DA could not
abolish the DA agonist induced yawning and
suppression of exploration. Thus, amphetamine
(0.2 mg/kg) pretreatment did not abolish the
behavioural effects of APO or pergolide in spite
of the fact that this dose of amphetamine
elevated extracellular DA when given in
conjunction with pergolide.
3. There was a poor correlation in time
between the effects of APO (0.05 mg/kg),
pergolide (0.02 mg/kg), EMD 23448 (2 mg/kg),
ocMPT (200 mg/kg) or reserpine (2 mg/kg) on
behaviour and reduction of DA levels.
APO-induced yawning and suppression of
exploration and pergolide- and EMD 23448-induced
yawning had a shorter latency than the reduction
of DA levels. The opposite relation was observed
for aMPT- or reserpineinduced suppression of
exploration.
4. Combined treatment with aMPT and APO, but
not pergolide, caused a more pronounced
suppression of exploration than did APO and
pergolide alone. The dose-response curve for
APO-induced yawning was not shifted by MPT (200
mg/kg) pretreatment.
Thus, the majority of the predictions made by
the autoreceptor hypothesis did not hold. The
only two findings supporting the autoreceptor
hypothesis are that reserpine suppressed
exploration and that the suppressive effect of
MPT on exploration added to that of APO
(however, not to that of pergolide).
It may be argued that the brain regions
investigated by microdialysis are not those
mediating yawning and suppression of
exploration. In fact, there is a considerable
amount of data supporting the hypothesis that
suppression of exploration by DA agonists can be
elicited from the accumbens. Given that these
findings can be taken as evidence that
suppression of exploration induced by
systemically administered
APO is also mediated by DA receptor
stimulation in the accumbens it would seem more
relevant to study the effects of APO and aMPT in
this region. We have found that the effect of
APO (0.05 mg/kg) on extracellular DA is smaller
in the accumbens (reduction to 80%) compared to
the striatum while the effect of ocMPT (50 mg/kg
causes reduction to 40%) is more similar to the
effect in the striatum.
DA agonist-induced yawning is probably
elicited from inside the blood-brain barrier.
However, local injections of DA agonists have
been found to elicit yawning in several
locations in the brain such as the striatum, the
septum, the central amygdaloid nucleus and the
hypothalamus. Lesion experiments have suggested
that the striatum is critically involved. Thus,
no definite conclusion can be drawn with respect
to the site (-s?) in the brain responsible for
mediating DA agonist-induced yawning. However,
the finding that combined treatment with
amphetamine and SCH 23390 induces yawning is a
very strong argument against autoreceptor
mediation, since it appears impossible that
post-synaptic D2-receptor stimulation can be
reduced by this treatment.
Thus, it is concluded that neither the
primary evidence in the literature, nor the
present findings constitute convincing evidence
in favour of the autoreceptor hypothesis. In
most cases, predictions made from the hypothesis
do not hold when tested. An alternative
hypothesis is that postsynaptic receptors
mediate yawning and suppression of exploration.
Another hypothesis for the mode of action of DA
agonists on behaviour must therefore be
proposed. The obvious alternative is that
populations of sensitive postsynaptic receptors
mediate DA agonistinduced yawning and
suppression of exploration. There is, however,
little direct evidence in favour of this
hypothesis. The most direct evidence is that
amphetamine plus SCH23390 induces yawning in a
dose where amphetamine given alone induces
strong stereotypies and causes an approximately
10-fold increase of the extracellular levels of
DA. The postsynaptic receptor hypothesis is
based partly on the assumption that there is a
variation in the sensitivity of DA receptors
between brain regions. The fact that behavioural
and electrophysiological effects of DA agonists
can be observed in different dose ranges
directly supports the validity of this
assumption. An important factor contributing to
the understanding of the variation in
sensitivity of different receptor populations is
the possibility of a difference in receptor
reserve between brain regions, between
postsynaptic receptors and autoreceptors
etc.
An important question is whether yawning and
suppression of exploration are equivalent or
not. Evidence that yawning and suppression of
exploration are pharmacologically distinct is
provided by the finding that scopolamine does
not antagonise suppression of exploration while
yawning is blocked by anticholinergics.
Furthermore, remoxipride inhibits APO induced
yawning but not suppression of exploration.
Thus, we conclude that yawning and suppression
of exploration are functionally distinct.
Clinical implications
One of the motives behind the massive amount
of research around the effects of low doses of
DA agonists is a number of reports that
DA-agonists are effective in various psychiatric
conditions such as schizophrenia and Gille
delaTourette's disease as well as neurological
disorders such as Huntington's chorea, tardive
dyskinesia and torticollis. Later reports have
been less optimistic, showing that the effects
on schizophrenia are shortlived and prone to
desensitization. The clinical efficacy of DA
agonists is a problem whose understanding
depends to some extent on the understanding of
behavioural models. ln view of the present data,
it is possible that the clinical effects of DA
agonists are not mediated by stimulation of DA
autoreceptors. In fact, it has been suggested
that the low dose effects of APO, because they
are selectively antagonised by sulpiride, may
serve as a model for negative symptoms of
schizophrenia. We note that the theoretical
understanding of the effects of DA agonists in
clinical states and animal models is a
completely different problem from the
demonstration of true clinical efficacy.