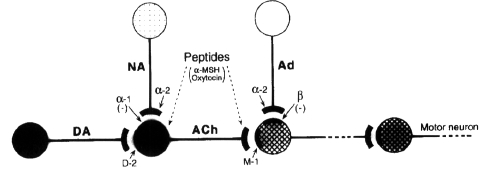
The administration of sinall doses of
apomorphine and other dopamine (DA) receptor
agonists produces repeated episodes of yawning
in rats (Mogilnicka
and Klimek 1977; Baggio and Ferrari 1983;
Protais et al. 1983; Serra et al. 1983a, b,
1984a, b, 1986a; Stahle and Ungerstedt 1984) and
in humans (Corsini et al. 1977).
This behavioural response scems to be due to
the stimulation of CNS DA receptors, being
prevented by centrally acting neuroleptics
(Mogilnicka and Klimek 1977; Nickolson and
Berendsen 1980; Dubec et al. 1982; Gower et al.
1984; Stahle and Ungerstedt 1984; Serra et al.
1986a) but not by domperidone (Gower et al.
1984; Stahle and Ungerstedt 1984, Serra et al.
1986 a), a DA receptor blocker which does not
cross the blood-brain barrier (Laduron and
Leysen 1979).
It has been suggested that DA receptors
mediating yawning might be identified with DA
autoreceptors, the stimulation of which results
in the inhibition of DA synthesis and release
and of dopaminergic firing (see Di Chiara et al.
1978). Consequently, yawning has been considered
a behavioural correlate of the inhibition of DA
transmission. However, this hypothesis is mainly
based on the fact that the doses of DA receptor
agonists needed to produce yawning are within
the same minute doses needed to stimulate DA
autoreceptors, while higher doses, which
stimulate both auto- and postsynaptic DA
receptors, produce locomotor hyperactivity and
stereotypy and suppress yawning (Mogilnicka and
Klimek 1977; Baggio and Ferrari 1983; Protais et
al. 1983; Stahle and Ungerstedt 1984; Gower et
al. 1984).
We have, however, recently provided evidence
suggesting that DA receptors mediating yawning
belong to a special population of central DA
receptors which are sensitive to DA agonists as
DA autoreceptors, but which are located
postsynaptically (Serra et al. 1986a).
The aim of the present study was to clarify
whether DA receptors mediating yawning were of
D1 (positively linked to adenylate cyclase) (see
Stoof and Kebabian 1981) or D2 type (not linked
or negatively linked to adenylate cyclase) (see
Stoof and Kebabian 1981; Onali et al. 1984).
Therefore, we compared the ability of
apomorphine, a mixed D1-D2 agonist (Kebabian and
Calne 1979), LY 171555, a selective D2 agonist
(Stoof and Kebabian 1984; Plantjé et al.
1985), and SKF 38393, a selective D1 agonist
(Stoof and Kebabian 1981), to induce yawning in
rats. We also studied the effect of sulpiride, a
selective D2 antagonist (Spano et al. 1979), and
SCH 23390, a selective D1 antagonist (Hyttel
1983), on the yawning response to apomorphine
and LY 171555. [...]
Wa have confirmed that apomorphine, a mixed
D1-D2 receptor agonist, induces yawning, while
the selective D1 agonist, SKF 38393, is
inactive. Morever, we have shown that LY 17155,
a selective D2 agonist, also induces
yawning. These results suggest that DA
agonist are of the D2 type. In accordance with
this hypothesis, we found that yaning response
to either apomorphine or LY 171555 was
antaginized by very small doses of the selective
D2 antagonist, (-) sulpiride. Thus, it was
surprising to find that minute doses of the
selective D1 antagonist SCH 23390 prevented both
apomorphine- and LY 171555-induced yawning.
One possible explanation for this paradox is
that SCH 23390, in spite of its in vitro
specifity for D1receptors, might also block in
vivo D2 receptors. This possibility is unlikely,
however, since there is convincing evidence
that, within the dose range used in the present
work, SCH 23390 also selectively blocked the
D1receptors subtype in vivo (Stoof and Kebabian
1984). Indeed, SCH 23390 has a low antiemetic
and hyperprolactinemic potency, as well as a
poor ability to antagonize dopaminergic
inhibition of syrnpathetic outflow, all
considered pure D2 effects. Chronic treatment
with SCH 23390 at the dose range used in the
present experiments produces a selective
increase in the number of striatal D1receptors,
and fails to modify D2 receptors (Porceddu et
al. 1985). Doses of SCH 23390 up to 1 mg/kg fail
to block DA autoreceptors (Mereu et al. 1985;
Gessa et al. 1986), a special population of D2
receptors (Lehmann et al. 1983; Pinnock 1983;
Walters et al. 1986).
The antagonism of SCH 23390 to the yawning
response cannot be explained by its action on
any known non-dopaminergic system. Thus, SCH
23390 has very little affinity for different
non-dopaminergic receptors such as those for
noradrenaline, histamine, acetylcholine and
benzodiazepines (Hyttel 1983; Cross et al. 1983;
Cristensen et al. 1984). At the doses which
antagonize yawning, SCH 23390 has no effect on
the classical serotoninergic behavioural
syndrome induced by the 5-HT agonists (Pugh et
al. 1985). The latter observation is in
agreement with the modest affinity of SCH 23390
for 5-HT receptors (Hyttel 1983; Cross et al.
1983) and also suggests that, while low doses of
SCH 23390 antagonize the yawning response, they
do not cause a general sedative effect
depressing behaviour.
It has recently been reported that stereotypy
and stimulation of motor activity induced by the
administration of apomorphine or other DA
agonists, which are considered D2 mediated
phenomena, are also potently antagonized by SCH
23390 (Christensen et al. 1984; Mailman et al.
1984; Molloy and Waddington 1984; Gessa et al.
1985; Pugh et al. 1985). These observations have
led the above authors to propose the existence
of a functional link between some populations of
D1and D2 receptors, so that blockade of the
former produces a functional inactivation of the
latter (Christensen et al. 1984; Gessa et al.
1985; Molloy and Waddington 1984; Pugh et al.
1985). According to Pugh et al. (l 985), such a
functional link might operate in the same
synaptic membrane in such a way that the
blockade of D1receptors alters the affinity of
D2 receptors for the agonists. Alternatively,
SCH 23390 might block D1receptors in a
dopaminergic system which exert a modulatory
influence on those processes which are initiated
by D2 stimulation. These considerations suggests
that the blockade of yawning by SCH 23390,
induced by selective stimulation of D2
receptors, might be explained assuming that,
like D2 receptors mediating motor activity and
stereotypy, D2 receptors mediating yawning are
also fonctionally interconnected with
D1receptors.
The finding that SCH 23390 antagonizes DA
agonist induced yawning provides indirect
support to our hypothesis that DA receptors
mediating yawning are distinct from DA
autoreceptors (Serra et al. 1986a). In fact, SCH
23390 is unique among neuroleptics in that it
fails to block DA autoreceptors (Mereu et al.
1985; Carlson et al. 1986; Gessa et al.
1986).
In conclusion, our results suggest that DA
receptors mediating yawning are a population of
postsynaptic DA receptors of the D2 type which
are as sensitive to DA agonists as DA
autoreceptors. Like DA receptors, which mediate
the classical excitatory responses of motor
activity and stereotypy induced by DA agonists,
D2 receptors mediating yawning appear to be
functionally linked to D1 receptors.
These findings further support our hypothesis
that yawning is a behavioural correlate of
increased DA transmission and might be
considered a first degree of arousal, mediated
by activation of a most sensitive population of
postsynaptic DA receptors (Serra et al. 1984b,
1986a, b). Further activation of DA transmission
might progress to higher level of arousal,
locomotor activity and stereotypy. This
interpretation is consistent with several
observations which suggest that yawning may be
considered a behavioural response subserving
arousal (see Serra et al. 1986b).
Gessa GL
Stretchings and yawnings induced by
adrenocorticotrophic hormone Nature 23/07/1966;
5047; 426-427
Gessa GL
Stretching and yawning movements after
intracerebral injection of ACTH Revue Canadienne
Bioliologie; 1967, 26, 3, 229-236
Kimura H,
Yamada K, Nagashima M, Matsumoto S, Ishii Y,
Yoshida S, Fujii K, Furukawa T Role of
adrenergic neuronal activity in the yawning
induced by tacrine and NIK-247 in rats.
Pharmacol Biochem Behav 1992; 43; 4; 985-91
Kimura
H, Yamada K, Nagashima M, Furukawa T
Involvement of catecholamine receptor activities
in modulating the incidence of yawning in
rats.Pharmacol Biochem Behav 1996; 53; 4;
1017-21
Laing J, Ogilvie R
EEG correlats of yawning during sleep onset
Sleep Research 1988; 17; 98
Molgilnicka
E REM sleep deprivation changes behavioral
response to catecholaminergic and serotoninergic
receptor activation in rats Pharmacol Biochem
Behav 1981; 15; 1; 149-151
Morelli M
et al Antagonism of apomorphine-induced
yawning by SCH 23390: Evidence against the
autoreceptor hypothesis
Serra G
, Collu M and Gessa GL Yawning is elicited
by D2 dopamine agonists but is blocked by D1
antagonist Psychopharmacology 1987; 91;
330-337
Serra G,
Gessa GL Hypophysectomy prevents yawning and
penile erection but not hypomotility induced by
apomorphine Pharmacology Biochemistry &
Behavior 1983; 19; 917-919
Serra G
et al Cycloheximide prevents apomporphine
induced yawning, penile erection and genital
grooming in rats European Journal of
Pharmacology 1983; 86; 279-282
Stahle
L Do autoreceptors mediate dopamine
agonist-induced yawning and suppression of
exploration ? a critical review.
Psychopharmacology 1992; 106; 1-13
Neumann BG,
Troncone LR, Braz S, Tufik S Modifications
on dopaminergic and cholinergic systems induced
by the water tank technique: analysis through
yawning behavior Arch Int Pharmacodyn Ther 1990;
308; 32-8
Stahle
L Do autoreceptors mediate dopamine
agonist-induced yawning and suppression of
exploration ? a critical review.
Psychopharmacology 1992; 106; 1-13
Tufik
S Does REM sleep deprivation induce
subsensitivity of presynaptic dopamine or
postsynaptic acetylcholine receptors in the rat
brain? European Journal of Pharrnacology 1987;
140; 215-219
Tufik S
Effects of stress on drug induced yawning
Physiology & behavior 1995; 8; 1;
1881-18
-Fugikawa
M; Yamada K; Nagashima M; Furukawa T
Involvement of beta-adrenoreceptors in
regulation of the yawning induced by
neuropeptides; oxytocin and alpha-melanocytes
stimuling hormone in rats. Pharmacol Biochem
Behav 1995; 50; 339-343
-Furukawa
T Yawning behavior for preclinical drug
evaluation Meth Find Exp Clin Phamacol 1996; 18;
2; 141-155
-Kimura H;
Yamada K; Nagashima M; Matsumoto S Role of
adrenergic neuronal activity in the yawning
induced by tacrine and NIK-247 in rats.Pharmacol
Biochem Behav 1992; 43; 4; 985-91
-Kimura
H; Yamada K; Nagashima M; Furukawa T
Involvement of catecholamine receptor activities
in modulating the incidence of yawning in
rats.Pharmacol Biochem Behav 1996; 53(; 4;
1017-21
Ogura
H, Kosasa T, Kuriya Y, Yamanishi Y Central
and peripheral activity of cholinesterase
inhibitors as revealed by yawning and
fasciculation in rats. Eur J Pharmacol. 2001;
415; 2-3; 157-64
-Serra
G , Collu M and Gessa GL Yawning is elicited
by D2 dopamine agonists but is blocked by D1
antagonist Psychopharmacology 1987; 91;
330-337
-Serra G,
Gessa GL Hypophysectomy prevents yawning and
penile erection but not hypomotility induced by
apomorphine Pharmacology Biochemistry &
Behavior 1983; 19; 917-919
-Serra G
et al Cycloheximide prevents apomporphine
induced yawning, penile erection and genital
grooming in rats European Journal of
Pharmacology1983; 86; 279-282
-Kostrzewa RM
and R Brus Is dopamine-agonist induced
yawning behavior a D3 mediated event? Life Sci
1991; 48; 26; 129
-Ushijima I,
Mizuki Y, Yamada M Multifocal sites of
action involved in dopaminergic-cholinergic
neuronal interactions in yawning
Psychopharmacology (Berl) 1988; 95; 34-7
-Ushijima
I et al, Muscarinic and nicotinic effects on
yawning and tongue protruding in the rat
Pharmacol Biochem Behavior 1984; 21;
297-300
-Ushijima
et al modification of apomorphine,
physiostigmine and pilocarpine induced yawning
after long term treatment with neuroleptic or
cholinergic agents Arch Int Pharmacodyn 1984;
271; 180-188
-Ushijima
I et al Characteristics of yawning behavior
induced by apomorphine, physostigmine and
pilocarpine Arch Int Pharmacodyn 1985; 273;
196-201
-Ushijima,
I., Y. Mizuki, et al. Behavioral effects of
dilazep on cholinergic, dopaminergic, and
purinergic systems in the rat. Pharmacol Biochem
Behav 1992;43(3): 673-676.
-Yamada K,
Furukawa T Direct evidence for involvement
of dopaminergic inhibition and cholinergic
activation in yawning Psychopharmacology 1980;
67; 39-43
-Yamada K,
Furukawa T The yawning elicited by
alpha-melanocyte-stimulating hormone involves
serotonergic -dopaminergic - cholinergic neuron
link in rats Naunyn-Schmiedeberg's Arch
Pharmacol 1981; 316; 155 -160
-Yamada K et
al Involvement of septal and striatal
dopamine D2 receptors in yawning behavior in
rats Psychopharmacology 1986; 90; 9-13
-Yamada K
et al Possible involvement of differing
classes of dopamine d2 receptors in yawning and
stereotypy in rats Psychopharmacology 1990; 100;
141-144
-Yamada K,
Furukawa T Behavioral studies on central
dopaminergic neurons. especially jumping,
stretching, body shaking and yawning behavior J
PharmacoBio dynamics 1980; 3; S16-S18