We have previously
demonstrated that the adrenergic mechanism seems
to be participated in inhibiting occurrence of
yawning because beta-adrenoceptor blockades and
inhibition of central adrenaline synthesis
caused by administration of synthesis enzyme
inhibitor similarly facilitate the occurrence of
yawning induced by dopaminergic and cholinergic
agonists.
On the other hand, the noradrenergic
mechanism seems to play a very important role in
modulating blood pressure responses, flexor
reflex activity, locomotor bchavior,
postdecapitation convulsions, and drinking
behavior. Regarding behavior, ambulation and
rearing were similarly decreased by an
alpha2-adrenoceptor agonist, clonidine,
alpha2-adrenoceptor antagonists, yohimbine,
rauwolscine, as well as piperoxan, and an
alpha1-adrenoceptor antagonist, prazosin.
However, grooming was decreased by clonidine and
prazosin but was markedly increased by
alpha2-adrenoceptor antagonists. As for yawning
behavior, a few reports have only been presented
with different results. SND 919, a dopamine
D2-receptor agonist, derived from an
alpha2-adrenoceptor agonist, clonidine, produced
stretching-yawning behavior that was reduced not
only by sulpiride but also by
alpha2-adrenoceptor antagonists, yohimbine and
idazoxan. An alpha -adrenoceptor antagonist,
prazosin, was reported to not to affect the
yawning induced by apomorphine or
antidepressants. However, Delini-Stula and Hunn
later proposed that the apomorphine-induced
yawning is potentiated by alpha1-adrenoceptor
antagonists and is suppressed by
alpha1-adrenoreceptor agonist. These
controversial findings have been of interest in
view of possible participation of
cathecholaminergic mechanism in modulating the
yawning. The present experiment were, therefore,
performed to investigate wheteher catecholmaine
recpetor activity is involced in regulating the
occurence of yawning. [...]
Discussion :
Previous experiments have shown that the
yawning induced by dopamine D2 receptor agonists
is antagonized by both dopamine and muscarine
receptor antagonists, and that by cholinesterase
inhibitors and muscarine receptor agonists is
inhibited by muscarine receptor antagonists, but
not by dopamine receptor antagonists. These
findings imply that the yawning induced by
dopamine receptor agonists involves activation
of dopaminergic and cholinergic neuronal
mechanisms and that by cholinesterase
inhibitors and direct muscarinic receptor
agonists involves activation of cholinergic
neuronal mechanisms. Consequently, it has been
proposed that the
dopaminergic-cholinergic neuronal link is
essentially participated in the occurrence of
yawning behavior.
In our
previous reports the yawning responses to
dopaminergic and cholinergic agonists were
increased by pretreatment with various
beta-adrenoceptor antagonists, such as pindolol,
propranolol, and indenolol, which block central
beta adrenoceptors after reaching the brain
through the blood-brain barrier, but not by
peripheral beta-adrenoceptor antagonists,
carteolol and atenolol. We have also found that
administration of phenylethanolamine,
N-methyltransferase inhibitors, which reduce
central adrenaline synthesis without affecting
noradrenaline levels, similarly increases the
yawning induced by tacrine, a cholinesterase
inhibitor, thus suggesting that the occurrence
of yawning evoked by both dopaminergic and
cholinergic activation is downregulated by
activity of central adrenergic neurons via
a-adrenoceptor stimulation. Indeed, the present
experiment also confirmed that pindolol
increased the yawning induced by talipexole,
physostigmine, and pilocarpine.
The discovery of presynaptic
alpha2-adrenoceptors, which play a role in
inhibition of noradrenaline release from
noradrenergic neurons, has permitted
subclassification of a-adrenoceptors into
alpha1- and alpha2-subtypes. As to
alpha1-adrenoceptors, there have been reports
describing that the yawning induced by
talipexole and apomorphine was unaffected by
prazosin at a lower dose of 1 mg/kg, IP, whereas
that by apomorphine was potentiated by prazosin
at a higher dose of 2.5 mg/kg, IP. In this
study, we used prazosin, 1-8 mg/kg, IP, and
bunazosin, 2.5-20 mg/kg, IP, at wide range of
dose, and the yawning produced by a dopamine
D2-receptor agonist, talipexole, was increased
after treatment with a,adrenoceptor antagonists,
such as prazosin (2-4 mg/kg) and bunazosin (5-
10 mg/kg), while the yawning induced by
physostigmine and pilocarpine was not increased
by the antagonists. Moreover, the
alpha1-adrenoceptor agonist ST587 markedly
suppressed the yawning induced by the
dopaminergic agonists, but not by the
cholinergic agonists. Consequently, the
noradrenergic neuronal mechanism seems to be
participated via alpha1-adrenoceptors in
decreasing the incidence of yawning caused by
the dopaminergic agonists, without influencing
the behavior induced by the cholinergic
agonists.
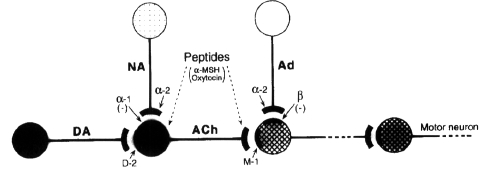
Interestingly, in the present experiment,
the beta-adrenoceptor antagonist, pindolol,
increased all the yawning responses to
talipexole, physostigmine, and pilocarpine,
whereas the alpha1-adrenoceptor antagonists and
agonists affected the responses to the
dopaminergic agonist, but not those to the
cholinesterase inhibitor and the direct
muscarinic receptor agonist. In addition, as
mentioned above, central adrenaline synthesis
inhibitors, which decrease adrenaline level
without changing noradrenaline level, potentiate
the yawning elicited by a cholinesterase
inhibitor. Therefore, it is presumed that, in
the dopaminergic-cholinergic neuronal link,
which is involved in causing the yawning
behavior, the noradrenergic mechanism inhibits
incidence of the behavior via
alpha1-adrenoceptor activity by interacting with
dopaminergic neurons that precede cholinergic
neurons, and the adrenergic mechanism inhibits
the incidence via beta-adrenoceptor activity by
interacting with cholinergic neurons.
As for alpha2-adrenoceptor antagonists, the
yawning evoked by talipexole was inhibited by
yohimbine and that by apomorphine and
physostigmine was reported to be reduced not
only by idazoxan and piperoxan, the
alpha2-adrenoceptor antagonists, but also by
clonidine, an alpha2-adrenoceptor agonist . In
the present experiment, selective
alpha2-adrenoceptor antagonists such as
idazoxan, rauwolscine, and yohimbine dose
dependently suppressed the yawning responses not
only to dopaminergic agonists but also to
cholinergic agonists. It has been proposed that
alpha2-adrenoceptors are located on
noradrenergic and/or adrenergic neuronal
pathways and that alpha2-adrenoceptor
antagonists increase both noradrenaline and
adrenaline release via blockade of
alpha2-adrenoceptors at central
catecholaminergic nerve terminals. Therefore,
although real mechanisms still remain uncertain,
the results may suggest a possibility that the
stimulation of noradrenergic and adrenergic
mechanisms elicited by an increase in
noradrenaline and adrenaline release induced by
alpha2-adrenoceptor blockade might result in
inhibition of the yawning evoked by both
dopaminergic and cholinergic activation.
In order to know more about the role of the
adrenoceptors on yawning from a physiological
point of view, it might be very important to
determine possible areas in the brain where the
yawning is involved, because of differential
distribution of adrenoceptors in the brain. We
have investigated that intracerebral injection
of dopamine receptor agonists into the striatum
and septum at low doses evoked yawning, but the
studies with other brain areas have not been
presented. Accordingly, this problem still
remain to be solved in future.
Although believed to be selective for a
specific receptor subtype, alpha-agonists and
antagonists act also on the other receptor
subtypes and/or receptors of other transmitters,
and the results, therefore, represent the
algebraic sum of their actions at the various
receptors involved. This will leave the
interpretation of the results speculative to a
certain extent until the neuronal pathways that
participate in the dopamine-acetylcholine link
in the control of the yawning response are well
identified.
In summary, the present findings suggest
that the catecholamine receptor activity is
involved in modulating the occurrence of yawning
in different manner.
Gessa GL
Stretchings and yawnings induced by
adrenocorticotrophic hormone Nature 23/07/1966;
5047; 426-427
Gessa GL
Stretching and yawning movements after
intracerebral injection of ACTH Revue Canadienne
Bioliologie; 1967, 26, 3, 229-236
Kimura H,
Yamada K, Nagashima M, Matsumoto S, Ishii Y,
Yoshida S, Fujii K, Furukawa T Role of
adrenergic neuronal activity in the yawning
induced by tacrine and NIK-247 in rats.
Pharmacol Biochem Behav 1992; 43; 4; 985-91
Kimura
H, Yamada K, Nagashima M, Furukawa T
Involvement of catecholamine receptor activities
in modulating the incidence of yawning in
rats.Pharmacol Biochem Behav 1996; 53; 4;
1017-21
Laing J, Ogilvie R
EEG correlats of yawning during sleep onset
Sleep Research 1988; 17; 98
Molgilnicka
E REM sleep deprivation changes behavioral
response to catecholaminergic and serotoninergic
receptor activation in rats Pharmacol Biochem
Behav 1981; 15; 1; 149-151
Serra G
, Collu M and Gessa GL Yawning is elicited
by D2 dopamine agonists but is blocked by D1
antagonist Psychopharmacology 1987; 91;
330-337
Serra G,
Gessa GL Hypophysectomy prevents yawning and
penile erection but not hypomotility induced by
apomorphine Pharmacology Biochemistry &
Behavior 1983; 19; 917-919
Serra G
et al Cycloheximide prevents apomporphine
induced yawning, penile erection and genital
grooming in rats European Journal of
Pharmacology 1983; 86; 279-282
Stahle
L Do autoreceptors mediate dopamine
agonist-induced yawning and suppression of
exploration ? a critical review.
Psychopharmacology 1992; 106; 1-13
Neumann BG,
Troncone LR, Braz S, Tufik S Modifications
on dopaminergic and cholinergic systems induced
by the water tank technique: analysis through
yawning behavior Arch Int Pharmacodyn Ther 1990;
308; 32-8
Stahle
L Do autoreceptors mediate dopamine
agonist-induced yawning and suppression of
exploration ? a critical review.
Psychopharmacology 1992; 106; 1-13
Tufik
S Does REM sleep deprivation induce
subsensitivity of presynaptic dopamine or
postsynaptic acetylcholine receptors in the rat
brain? European Journal of Pharrnacology 1987;
140; 215-219
Tufik S
Effects of stress on drug induced yawning
Physiology & behavior 1995; 8; 1;
1881-18
-Fugikawa
M; Yamada K; Nagashima M; Furukawa T
Involvement of beta-adrenoreceptors in
regulation of the yawning induced by
neuropeptides; oxytocin and alpha-melanocytes
stimuling hormone in rats. Pharmacol Biochem
Behav 1995; 50; 339-343
-Furukawa
T Yawning behavior for preclinical drug
evaluation Meth Find Exp Clin Phamacol 1996; 18;
2; 141-155
-Kimura H;
Yamada K; Nagashima M; Matsumoto S Role of
adrenergic neuronal activity in the yawning
induced by tacrine and NIK-247 in rats.Pharmacol
Biochem Behav 1992; 43; 4; 985-91
-Kimura
H; Yamada K; Nagashima M; Furukawa T
Involvement of catecholamine receptor activities
in modulating the incidence of yawning in
rats.Pharmacol Biochem Behav 1996; 53(; 4;
1017-21
Ogura
H, Kosasa T, Kuriya Y, Yamanishi Y Central
and peripheral activity of cholinesterase
inhibitors as revealed by yawning and
fasciculation in rats. Eur J Pharmacol. 2001;
415; 2-3; 157-64
-Serra
G , Collu M and Gessa GL Yawning is elicited
by D2 dopamine agonists but is blocked by D1
antagonist Psychopharmacology 1987; 91;
330-337
-Serra G,
Gessa GL Hypophysectomy prevents yawning and
penile erection but not hypomotility induced by
apomorphine Pharmacology Biochemistry &
Behavior 1983; 19; 917-919
-Serra G
et al Cycloheximide prevents apomporphine
induced yawning, penile erection and genital
grooming in rats European Journal of
Pharmacology1983; 86; 279-282
-Kostrzewa RM
and R Brus Is dopamine-agonist induced
yawning behavior a D3 mediated event? Life Sci
1991; 48; 26; 129
-Ushijima I,
Mizuki Y, Yamada M Multifocal sites of
action involved in dopaminergic-cholinergic
neuronal interactions in yawning
Psychopharmacology (Berl) 1988; 95; 34-7
-Ushijima
I et al, Muscarinic and nicotinic effects on
yawning and tongue protruding in the rat
Pharmacol Biochem Behavior 1984; 21;
297-300
-Ushijima
et al modification of apomorphine,
physiostigmine and pilocarpine induced yawning
after long term treatment with neuroleptic or
cholinergic agents Arch Int Pharmacodyn 1984;
271; 180-188
-Ushijima
I et al Characteristics of yawning behavior
induced by apomorphine, physostigmine and
pilocarpine Arch Int Pharmacodyn 1985; 273;
196-201
-Ushijima,
I., Y. Mizuki, et al. Behavioral effects of
dilazep on cholinergic, dopaminergic, and
purinergic systems in the rat. Pharmacol Biochem
Behav 1992;43(3): 673-676.
-Yamada K,
Furukawa T Direct evidence for involvement
of dopaminergic inhibition and cholinergic
activation in yawning Psychopharmacology 1980;
67; 39-43
-Yamada K,
Furukawa T The yawning elicited by
alpha-melanocyte-stimulating hormone involves
serotonergic -dopaminergic - cholinergic neuron
link in rats Naunyn-Schmiedeberg's Arch
Pharmacol 1981; 316; 155 -160
-Yamada K et
al Involvement of septal and striatal
dopamine D2 receptors in yawning behavior in
rats Psychopharmacology 1986; 90; 9-13
-Yamada K
et al Possible involvement of differing
classes of dopamine d2 receptors in yawning and
stereotypy in rats Psychopharmacology 1990; 100;
141-144
-Yamada K,
Furukawa T Behavioral studies on central
dopaminergic neurons. especially jumping,
stretching, body shaking and yawning behavior J
PharmacoBio dynamics 1980; 3; S16-S18