- 2006
Le
bâillement son histoire interne ou
neurophysiologie du
bâillement
- Electroneurobiologia
2006 in press
-
- « On a vainement
déroulé les plis du cerveau, ils
sont restés muets à cette
investigation; ils n'ont
révélé aucune trace des
actes sublimes qui s'y opèrent pendant la
vie. » Jean-Louis
Brachet. Physiologie
élémentaire de l'Homme,
Germer-Baillière Ed, 1855
-
- Trois comportements rythment la vie des
vertébrés : l'éveil, le
sommeil profond et le sommeil paradoxal. Le
sommeil est caractérisé par une
posture, correspondant
généralement à la
relaxation des muscles antigravitaires et une
réduction des perceptions
environnementales. Il est régulé
par des processus homéostasiques
couplés à des rythmes circadiens
et ultradiens
-
- Les
différentes étapes de la
différenciation du cerveau chez les
vertébrés est à la base de
deux états d'éveil. Le plus ancien
est l'activité diurne, liée
à la lumière au sein d'un rythme
circadien. Les poïkilothermes
contrôlent cet état
entièrement à partir du tronc
cérébral, siège des aires
sensori-motrices. Les mammifères ont
développé les structures
thalamo-corticales qui ont déplacé
l'éveil aux structures corticales
simultanément à l'acquisition de
l'homéothermie et de la capacité
à une activité nocturne. Afin
d'éviter une duplication
compétitive des compétences de ces
deux structures, le type d'éveil le plus
ancien contrôlé par le tronc
cérébral est inhibé et
pourrait correspondre au sommeil lent (Nicolau,
2000). L'effet inducteur du sommeil propre
au noyau VLPO, parachiasmatique et donc
anatomiquement à la partie rostrale du
diencéphale, phylogenétiquement
plus récente, s'accorde avec ce statut
inhibiteur (GABA). La découverte chez les
monotrèmes, mammifères
protothériens (l'échnidé et
l'ornithorynque) d'un temps de sommeil paradoxal
supérieur à 60% du temps total de
sommeil, proportion la plus élevée
de tous les mammifères, conforte la loi
proposée par Ernst
Haeckel (1834-1919) : l'ontogenèse
reproduit la phylogénèse (Siegel,
1999). Forme ontogénique la plus
précoce de sommeil, le sommeil paradoxal
a une origine rhombencéphalique caudale
(tronc cérébral inférieur,
cholinergique, "archaïque"), source de
l'inhibition motrice descendante spinale.
L'instabilité de la régulation
thermique (type poïkilotherme) s'ajoute
à l'origine phylogénétique
plus ancienne pour proposer l'analogie entre le
sommeil paradoxal et l'état inactif
nocturne des reptiles (archéosommeil).
Enfin, il faut noter l'évolution de la
proportion des différents types
d'état de vigilance au cours du
nycthémère, chez l'homme, qui
montre, de la vie intra-utérine à
la vieillesse, une augmentation de la
durée d'éveil actif, une
réduction du temps global de sommeil avec
réduction prédominante du sommeil
paradoxal (Chalamel, 1992 ; Siegel, 2005 ;
Valatx, 2004).
-
- Le
bâillement, qu'on peut
interpréter comme une pandiculation
partielle ne touchant que les muscles de la
face, du cou et du diaphragme, est un
comportement contemporain des transitions entre
ces états.
Phylogénétiquement très
ancien, puisque déjà
présent chez les reptiles, il persiste
inchangé jusqu'aux primates humains et
peut s'observer dans les mondes aériens
et aquatiques. Ses moments
privilégiés d'apparition sont
l'éveil ou lors du besoin de sommeil
avant l'endormissement. Sauf pathologie chez
l'homme, il n'apparaît jamais pendant le
sommeil. Le bâillement est un comportement
ontogenétiquement très
précoce, apparaissant dès la
12° semaine de grossesse. Le nombre
quotidien de bâillements évolue au
cours de la vie. On peut constater une
réduction progressive du nombre de
bâillements quotidiens de façon
parallèle à la réduction de
la proportion de sommeil paradoxal, de la vie
intra-utérine à la vieillesse. Les
mammifères ayant la plus grande
immaturité cérébrale
à la naissance, c'est à dire pour
qui la myélinisation caudo-rostrale de
leur système nerveux doit s'achever
après la naissance, sont ceux ayant la
plus grande part de sommeil paradoxal à
la naissance, et dont la proportion (REM - non
REM sleep) se réduit nettement au cours
de la vie. L'observation comportementale des
prématurés humains, grands
bâilleurs, indique là aussi un
parallèle entre nombre de
bâillements et importance de la proportion
de sommeil paradoxal (Giganti,
2002).
-
- J.
Siegel (1999), qui a mis en évidence
l'existence d'un sommeil paradoxal
prolongé des monotrèmes,
n'a jamais constaté, chez eux, de
comportement pouvant ressembler à un
bâillement. Ce témoignage,
basé sur des souvenirs, car l'observation
des bâillements ne faisait pas partie du
programme de recherche, incite à proposer
une réelle étude, à
prolonger chez les marsupiaux (mammifères
protothériens) pour lesquels n'existent
aucune donnée concernant l'existence de
bâillements.
-
- La description classique du bâillement
le présente comme un cycle respiratoire
paroxystique. Il paraît possible de
proposer une autre théorie. En
réexaminant la pandiculation dans son
entier déroulement, il est possible
d'interpréter ce comportement comme un
travail musculaire
généralisé avec :
- Ð une contraction des muscles de la
mâchoire et du pharyngo-larynx
(diamètre multiplié par 4, ce qui
ne se retrouve dans aucun autre comportement
physiologique), responsable de la large
ouverture de bouche et des voies respiratoires
hautes,
- Ð une contraction massive du diaphragme
responsable de l'ample inspiration,
- Ð une contraction des muscles extenseurs
des membres et des muscles paravertèbraux
responsable de l'étirement des membres et
du tronc.
-
- Il existe une tendance à
l'augmentation progressive de l'activité
vagale, de la veille au sommeil lent, avec
pic lors du sommeil paradoxal tonique tandis que
l'activité sympathique
décroît presque
parallèlement. À l'état de
veille, la dépression engendrée
par l'inspiration provoque une constriction des
voies respiratoires qui est physiologiquement
contre-balancée par une contraction
active des dilatateurs pharyngolaryngés
(muscles génioglosses,
génohyoïdiens) (Ayappa,
2003)
-
- Les expériences
électrophysiologiques (stimulation du
locus cœruleus) et pharmacologiques ont
montré un effet facilitant
noradrénergique sur les motoneurones
spinaux, source de l'activité musculaire.
La suspension de l'activité tonique du
locus cœruleus adrénergique et
d'autres noyaux du pont, lors du sommeil
paradoxal, est responsable de l'hypotonie
périphérique (Pace-Schott,
2002). Il en résulte,
également, une diminution de
l'activité des muscles dilatateurs des
voies aériennes supérieures,
expliquant une tendance au collapsus du
pharyngo-larynx. Celle-ci est maximale lors du
sommeil paradoxal, plus prolongé en
Þn de nuit. Lors de l'éveil, la
reprise de l'activité
noradrénergique déclenche une
récupération de l'activité
motrice active. L'étirement musculaire
des membres et du tronc témoignent de la
reprise musculaire tonique déclenchant
une augmentation de la fréquence
cardiaque, de la pression artérielle, du
métabolisme musculaire associée
à un dérouillage articulaire. Le
bâillement apparaît comme une ample
contraction des muscles inhibés lors du
sommeil paradoxal, un peu comme un effet en
miroir inverse de l'inhibition provoquée
par le sommeil paradoxal, ouvrant largement le
pharyngo-larynx. Si les motoneurones
stimulés sont sus-jacents à la
V° racine cervicale, un bâillement
apparaît isolément, comportant les
contractions-étirements des muscles de la
face, du cou et du diaphragme (nerf
phrénique). Si l'ensemble des noyaux
moteurs du tronc cérébral et des
voies spinales est stimulé, la
pandiculation apparaît (Beyaert,
1998).
-
- La suppression expérimentale du
sommeil paradoxal s'est accompagnée d'une
disparition des bâillements. Menées
chez le rat de laboratoire Sprague Dawley, ces
expériences induisent un stress
empêchant le passage en sommeil pardoxal.
Ce stress provoque une stimulation de la
sécrétion cortisolique, produit
Þnal de l'activation de l'axe
hypothalamo-hypophyso-surrénalien. Cet
hypercorticisme induit, altèrerait
l'activité dopamino-cholinergique
responsable du bâillement. Mais, en
réalité, est-ce le stress ou
est-ce l'absence de sommeil paradoxal qui fait
disparaître les bâillements
(Lobo, 1990
; Molgilnicka,
1981 ; Neumann,
1990 ; Tufik,
1987) ?
-
- Il paraitrait intéressant
d'approfondir une recherche envisageant les
liens ontogéniques,
phylogénétiques et comportementaux
unissant sommeil paradoxal et
bâillements.
-
- Flip-flop switch et
bâillements
-
- Walter
Cannon (1871-1945) a défini
l'homéostasie comme
« l'ensemble des processus organiques
qui agissent pour maintenir l'état
stationnaire de l'organisme, dans sa morphologie
et dans ses conditions intérieures, en
dépit de perturbations
extérieures » (Fleming, 1984).
Or, l'état naturel du monde vivant est le
manque. Les mécanismes
homéostasiques physiologiques,
régulés par l'hypothalamus, sont
adaptés à cette gestion du manque
expliquant ainsi l'asymétrie des
mécanismes neuro-hormonaux mis en oeuvre.
Pour chacun existe une redondance des
mécanisme stimulants (éveil) et
d'épargne (énergie,
équilibre hydro-électrolytique)
par rapport aux mécanismes de
dépense et d'élimination le plus
souvent unicistes (insuline, peptides
natriurétiques). Bâillements et
pandiculations semblent apparaître lors de
l'action des systèmes
d'homéostasie stimulants (éveil,
sexualité) et d'épargne (jeune,
hypoglycémie, hypoTA) (Walusinski,
2004).
-
- La
régulation des états de veille et
de sommeil sont profondémment
intégrés aux systèmes
d'homéostasie, régulés par
l'hypothalamus comme le sont l'appétit,
la température corporelle et le
métabolisme. Plusieurs mécanismes
redondants maintiennent l'éveil,
état indispensable à la survie;
seul le noyau VLPO assure l'endormissement.
-
- Le bâillement
peut-il représenter un comportement
observable, témoin des mécanismes
neurobiologiques faisant basculer de
l'éveil au sommeil et
réciproquement?
-
- Pendant et après la fin de la
première guerre mondiale (1918-1926), une
pandémie présumée d'origine
vrale, la célèbre grippe
espagnole, coïncida avec une
épidémie d'encéphalites
laissant nombre de victimes dans des
états prolongés d'endormissement
ou léthargie compliqués
secondairement de syndromes extra-pyramidaux;
paradoxalement quelques victimes avaient, elles,
une insomnie rebelle et des mouvements
choréiques.
-
- On doit au neurologue viennois, le Baron
Constantin von
Economo, la description de cette pathologie
et de la localisation anatomopathologique des
troubles. Dès ce travail princeps,
l'absence de contagiosité directe, de
mise en évidence d'un agent infectieux
avait fait douter du lien entre grippe et
encéphalite. Il est actuellement admis
une origine auto-immune, rappelant ainsi les
mécanismes responsables de la
narcolepsie, peut-être
post-streptococcique. Les survivants ont souvent
manifestés de séquelles chroniques
à type d eparkinsonnisme, de dystonies.
Mais pour notre propos, notons l'association des
troubles du sommeil et de troubles
neuro-endocriniens (diabète insipide,
obésité, aménorrhée,
impuissance...)
-
- Von Economo remarqua chez
les léthargiques des lésions
neuronales de l'hypothalamus postérieur
et du mésencéphale, partie
rostrale du tronc cérébral; chez
les insomniaques, les lésions
siègeaient dans l'aire préoptique
et le diencéphale. Fort de ses
constations, von Economo prédit que les
régions hypothalamiques proches du
chiasma optique contenaient les neurones
responsables de l'endormisssement, alors que
ceux de l'hypothalamus postérieur
étaient responsables de
l'éveil.
-
- La véracité de ces
prédictions s'est vue peu à peu
confirmée. Le noyau supra-chiasmatique et
l'hypothalamus ont été
identifiés comme responsables des rythmes
circadiens. Si la découverte de la
réticulée ascendante aminergique
(diencéphale et
mésencéphale) responsable de
l'éveil (Moruzzi et Magoun) a, pour un
temps déplacé
l'intérêt pour les structures
éveillantes vers le tronc
cérébral, la mise en
évidence de l'hypocrétine,
neurotransmetteur hypothalamique indispensable
au maintien de l'éveil,
sécrété dans l'hypothalamus
latéral (de Lecea, Sakurai), a
réactualisé l'hypothèse de
von
Economo.
-
- Le noyau ventrolatéral
préoptique (VLPO) a, lui, une action
inhibitrice GABA dirigée sur les noyaux
tubéro mammillaires(histaminertgiques),
les noyaux du raphé
(sérotoninergiques) et le locus coeruleus
(aminergique), toutes structures responsables de
l'éveil; il apparaît donc comme une
structure majeure de l'endormissement (Sherin,
Saper, Luppi).
-
- De façon schématique, notre
état balance d'un état
d'éveil à un état de
sommeil et l'inverse, par une sorte d'inhibition
réciproque. L'électronique
connaît ce type d'interrupteurs bistables,
baptisés flip-flop switch. On
conçoit ainsi un passage rapide d'un
état à l'autre, condition
essentielle de survie du monde animal.
-
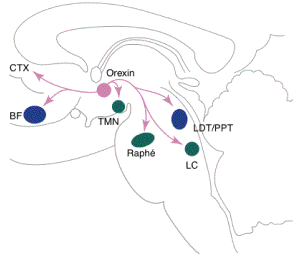
- L'hypocrétine
apparaît comme indispensable au maintien
de l'éveil avec tonus musculaire
adapté. Son déficit est
responsable de la catalepsie de la narcolepsie.
Les neurones à hypocrétine
semblent jouer le rôle du doigt qui appuie
sur l'interrupteur flip-flop switch, maintenant
l'éveil.
-
- Le bâillement peut se concevoir comme
un comportement n'apparaîssant que lors
des transitions entre les deux états
stables éveil / sommeil, seul
témoin comportemental observable des
changements d'état du flip-flop
switch.
-
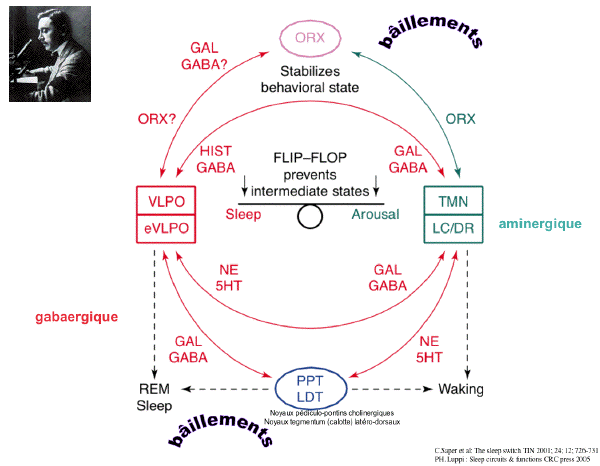 -
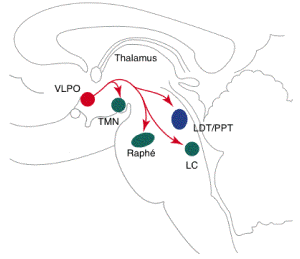
- A model for reciprocal interactions between
sleep and wakepromoting brain regions, which
produces a flipÐflop switch.
- inhibitory pathways are
shown in red
- the excitatory
pathways in
green
- the blue circle
indicates neurons of the LDT and PPT
- green boxes
indicate aminergic nucle
- the red box
indicates the VLPO
- Aminergic regions such as the TMN, LC and DR
promote wakefulness by direct excitatory effects
on the cortex and by inhibition of
sleep-promoting neurons of the VLPO.
- During sleep, the VLPO inhibits
amine-mediated arousal regions through GABAergic
and galaninergic (GAL)projections.
- Most innervation of the TMN originates in
the VLPO core, and input to the LC and DR
predominantly comes from the extended VLPO. This
inhibition of the amine-mediated arousal system
disinhibits VLPO neurons, further stabilizing
the production of sleep. The PPT and LDT also
contain REM-promoting cholinergic neurons.
- The extended VLPO (eVLPO) might promote REM
sleep by disinhibiting the PPT-LDT; its axons
innervate interneurons within the PPT-LDT, as
well as aminergic neurons that normally inhibit
REM-promoting cells in the PPT-LDT.
Orexin/hypocretin neurons (ORX) in the lateral
hypothalamic area (LHA) might further stabilize
behavioral state by increasing the activity of
aminergic neurons, thus maintaining consistent
inhibition of sleep-promoting neurons in the
VLPO and REM-promoting neurons in the
PPT-LDT.
- Unbroken lines represent neuronal pathways
described in the text.
- Broken black lines indicate influences of
specific regions on behavioral states.
-
- Abbreviations:
- DR, dorsal raphé nucleus
- HIST, histamine
- LC, locus coeruleus
- LDT, laterodorsal tegmental nuclei
- PPT, pedunculopontine tegmental nuclei
- REM, rapid eye movement
- TMN, tuberomammillary nucleus
- VLPO, ventrolateral preoptic nucleus
-
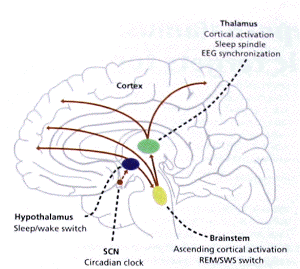
- Hypothalamic and brainstem sleep/wake
regulation systems, in relation to common sleep
disorders and their pharmacological treatment
(a) Distinct roles of the brainstem, thalamus,
hypothalamus and cortex for vigilance control.
(b)
-
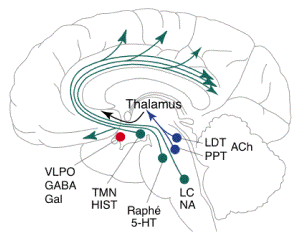
- Recent discoveries of the sleep-promoting
GABAergic/galanininergic (Gal) neurons in the
ventrolateral preoptic area (VLPO) and the
wake-promoting hypocretin/orexin neurons in the
lateral hypothalamus focus attention on the
hypothalamus. Destruction of these systems
induces insomnia and narcolepsy, respectively,
mirroring the pioneering clinical observations
during the encephalitis lethargica epidemic.
ALThough no direct interaction between the two
systems has been reported, both VLPO and
hypocretin systems innervate the main components
of the ascending arousal system, such as
adrenergic locus coeruleus (LC), serotoninergic
dorsal raphe (DR) and histaminergic
tuberomammillai-y nucleus (TNIN).
-
- The VLPO system inhibits, and the hypocretin
system activates, these systems. Thus, the
hypothalamus may serve as a center for the
'sleep switch' under the influence of the
circadian clock.
-
- In addition to these ascending arousal
systems, pharmacological evidence suggests the
involvement of dopamine (DA), especially in the
ventral tegmental area (A 10), for the control
of alertness. The DA system, including
descending A11 projections, may also be
particularly important for sleep-related motor
control in conditions such as cataplexy and
PLM.
-
- Recent anatomical and pharmacological
evidence suggests that serotonin (5HT) is
important for the motor control of the
hypoglossal nucleus, and dysregulation of this
system may be involved in upper airway
resistance in obstructive sleep apnea.
-
- Histamine (His) is a wake-promoting amine,
but it has a variety of peripheral actions.
Histaminergic compounds are not widely used in
sleep medicine, except the CNS-permeable HI
antagonists for a small number of insomnia
patients.
-
- Benzodiazepines (BZs) act on the GABAA/BZ-Cr
macromolecular complex, and may act on any part
of the arousal system shown here.
-
- BF, basal forebrain cholinergic nuclei; LDT/
PPT, laterodorsal tegmental
nuclei/pedunculopontine tegmental nuclei; CR,
caudal raphe; PRF, pontine reticular formation;
ACh, acetylcholine; PLM, periodic leg movements
during sleep; NE, norepinephrine; GLY, glycine;
GLU, glutamate.
-
     -
|


