Why we
sleep : the evolutionary pathway to the
mammalian sleep
MC. Nicolau, M. Akaarir, A. Gamundi, J.
Gonzalez, RV. Rial
Laboratori de fisiologia,
Universitat des illes Balears, Palma de
Mallorca
[...] The phylogenetic appearance of
sleep can be approached through a study of the
evolution of the vertebrate brain. This began as
an undifferentiated dorsal nerve, which was
followed by the development of an anterior
simplified brain and ended with the formation of
the multilayered mammalian neocortex or the
avian neostriate.
The successive stages in the differentiation
of the vertebrate brain produced, at least, two
different waking types. The oldest one is the
diurnal activity, bound to the light phase of
the circadian cycle. Poikilotherms control the
waking from the whole brainstem, where their
main sensorymotor areas lie.
Mammals developed the thalamocortical lines,
which displaced the waking up to the cortex
after acquiring homeothermy and nocturnal
lifestyle. In order to avoid competence between
duplicate systems, the early waking type,
controlled from the brainstem, was suppressed,
and by necessity was turned into inactivity,
probably slow wave sleep. On the other hand, the
nocturnal rest of poikilotherms most probably
resulted in rapid eye movement sleep. The
complex structure of the mammalian sleep should
thus be considered an evolutionary remnant; the
true acquisition of mammals is the cortical
waking and not the sleep.
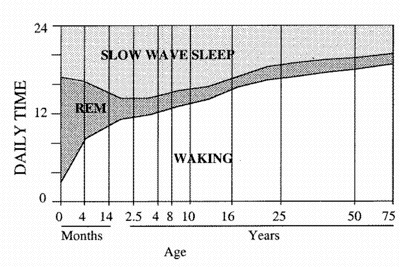
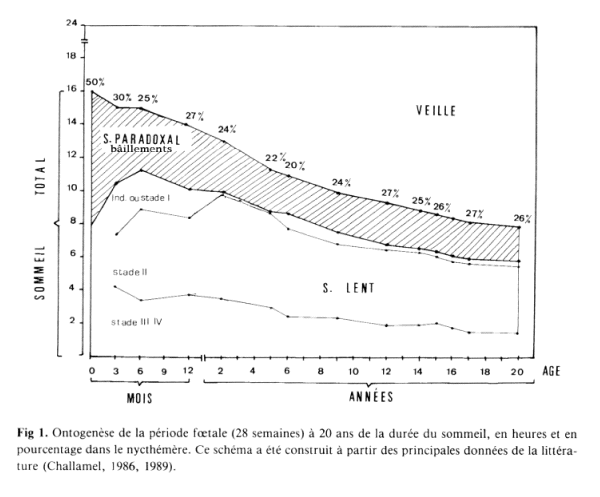
The relationships between REM, non-Rem (WSW)
and waking changes with age. Yawn's
frequencies and REM sleep evolve in
parallel.
The reptiles seern to be not too different from
the rest of poikilotherms, thus making
acceptable the search of traits of primitive
sleep in these animals. They have two types of
behavioral sleep: one nocturnal and associated
to the circadian cycles of external light and
heat, while the other is diurnal and
independent, at a first sight, of external
factors, although this really means that
eventual factors deterrnining it are not easily
observable. Surprisingly, these features show
their maximal expression in active animals and
not in behaviorally sleeping ones.
In relation to REM, the classical mammalian
indicators have not been clearly found in
reptiles, but the evidence reported in
monotremes seems to be definitive in suggesting
that REM is, at least, as old as SWS. Finally,
some evidence points to a homology between the
whole nocturnal sleep of reptiles and the
mammalian REM. [...]
The similarity between manimalian SWS and
reptilian waking is much clearer than the
supposedly obvious between the two waking
states. On the contrary, while achieving the
same purpose, i.e. maintaining behavioral
activity, the waking states are dissimilar in
both anatomic and neurological terms. This led
to assert the homology (common phylogenetic
origin with independence of function) between
mammalian SWS and reptilian waking.
One could arrive to the same conclusions in
which the main parts of the mammalian brain have
been displayed according to their hierarchical
relationship to the activity states. The cortex
is in the hierarchical top during waking. It
produces the most important part of the sensory
processing and the behavioral output, while the
remaining lower centers only provide support for
the necessary accompanying furictions, such as
auxiliary movements, homeostatic adjustments and
so on. The situation changes during SWS, because
the cortex remains inactivated during this state
due to diencephalic ascendant inhibition.
Therefore, the brainstem (we include the
diencephalon, mesencephalon and rombencephalon
under this name) lies in the top hierarchical
level during this state while the cortex is kept
at the bottom. Finally, there is an additional
hierarchical change during REM: now the
rombencephalon is on top. The cortex is in full
activity during REM but it remains disconnected
in sensory and motor terms due to rombencephalic
inhibitory influence. In consequence, it has
been placed second in the hierarchical control
during this state. Finally, the brainstern lies
in the lower hierarchical level during this
state, in correspondence with the low
homeostatic capacity of REM. The arrows
represent the directions of level change
corresponding to the transitions between waking,
SWS and REM. In addition to the transitions
represented, there is also the one from REM to
SWS or from SWS to waking, but not a direct
transition from waking to REM.
In consequence, reptiles cannot have a
waking of the same type than that of mammals
(cortical waking). This does not means that they
cannot be vigilant, but that their vigilance is
achieved using the same anatomical regions used
by mammals to be asleep in SWS. Deleting the
cortex causes no functional change; the
rombencephalic inhibition travelling downwards
would cause inactivity, both in mammals and in
submammals. In conclusion, the REM and the
inactivity of poikilotherms are both homologous
and analogous.
"Embryogenesis wifi repeai phylogenesis in
situations kvere embryologicalprocesses occur in
a causal-continuity because the evolutive
conditions for this sequence to be built must
have occurred through sequential continuous
modifications- (Horder, 1989). The main lines in
the embryogencsis of the nervous system clearly
repeats the phylogeny. In consequence, the
activity states should repeat the phylogeny, as
they maintain a causal continuity with the
anatomic structure of the brain. The sleep could
be thus a paradigmatic example in the
application of the embryogenetic law and the
particular features of the rnammalian
polygraphic sleep could be unavoidable results
of the evolutionary development of the brain. A
mammalian waking is only possible, when the
neocortex reaches maturity, and this occurs in
the last stages of the ontogeny, what was called
"the advanced wakefulness".
When the correlation between sleeping time
and brain development was recognized, the sleep
seemed needed for brain maturation. Many
experimental studies have found severe
impairments in brain structure and function
consequent to deprivation or reduction in the
normal amount of REM. But the relation could be
inverse: sleep might be the only possible state
during stages of low brain development. A
controversy between these two points of view is
asymmetrical in more than one aspect: first,
from a plain philosophical view point, the
simplest hypothesis should always be preferred,
and is simpler to suppose the unavoidability of
REM in a under developed brain in front of a
complex (and unknown) REM dependent mechanism of
brain arowth and differentiation.
Why an unavoidable bond between REM and
brain maturation should have developed? Second,
the need of REM for a correct development of the
brain has not been discussed in front of the
option of REM unavoidability due to low brain
capacity. Experimentation should be performed to
test the former option in a similar amount as
the one performed to demonstrate the latter.
Last, but not least, the experiments showing the
need of REM for brain development provide with
arguments of necessity but not of sufficiency.
On the contrary, an undeveloped brain is a
sufficient condition to produce a state of
reduced behavioral output, as one would qualify
the REM. It is a priori clear that a much
smaller amount of brain is needed to produce
rest, than needed to produce vigilance. However,
as total rest is practically impossible in live
beings, some kind of output should exist even in
an undeveloped brain and this necessary output
is what we call the REM signs.
According to the foregoing arguments, the
question of why we sleep has lost sense. Most
important than their sleep, mammals gained a
thick neocortex and a waking with unparalleled
features in the animal kingdom. We just sleep
while we are not cortically vigilant. The
complexity of the mammalian sleep is only a side
effect, which probably does not add too much to
the environmentally driven rest- activity cycles
of simpler animals. According to Williams rules,
the sleep does not need to be adaptive by itself
as it perfectly fits in the group of byproducts
of other truly adaptive changes.
[...]
"SWW turned SWS" hypothesis: first,
two changes are needed to explain the production
of waking, one for the cortical mammalian waking
and another one for the striatal waking of
birds. No change is necessary to explain the
production of SWS as it passed relatively
unchanged from being SWW to be transformed in
truc SWS (it is the result of the same change
which produced waking, it should not be taken
into account more than once). Finally, one
mutation is needed to explain the possible
disappearance of REM in the echidna. In summary,
only two, or three at most, changes are needed
to explain the evolution of sleep. However, all
are known for sure: the first two ones, needed
to explain the changes in the telencephalic
structure are undeniable. As it has been already
explained, the changes in waking and sleep would
only be their side results. The third, already
explained, is a side result of the speciation
process between platypus and echidnas. In
summary, the hypothesis defended in this review
needs two mutations, the same found for the
hypothesis "REM first" but it is fact more
parsimonious, because it explains, not only the
sleep, but also the waking, an important missing
aspect in the other two alternatives.
In addition to showing higher parsimony, the
"SWW turned SWS" hypothesis has other additional
virtues: we saw how it is well congruent with
the phylogenetic law, with the
altricial-precocial dimension and also explains
the thermogenic correlations of the waking-sleep
states. The alternative hypotheses however,
provide no reasons to explain why the supposed
phylogenetic changes took place. Those theories
meet perhaps the requisite of necessariness, but
provide no hint at all neither on sufficiency
nor on historical unavoidability. Unlike them,
for the "SWW turned SWS "theory, it was
necessary to produce deep changes in the
structure of the nervous system for a reptile to
get adapted to live in the dark and these
changes were reasonably sufficient to produce
the two types of mammalian sleep.
They were unavoidable to gain the nocturnal
niche, constrained by the earlier structure of
the brain; the only solution was to suppress
duplicated sensory and motor systems, but these
systems were intermingled with essential
homeostatie functions. Therefore, it was
impossible to produce their complete
annihilation. Achieving a selective block was
thus, once again, unavoidable. Some functions of
the brainstem had to be suppressed while others
should bc maintained. The result was the SWS, a
state of high brainstem activity and low
behavioral output. The organism ended with two
states of rest: the old one, which resulted in
the modern REM and the new one, the SWS. This
chain of events was necessary to cope with the
new demands of a dramatic change in the
lifestyle, but it was also, with high
verisimilitude, a sufficient cause. The
development of a new waking type is a sufficient
reason to always block any other less efficient
vigilance, i.e., the creation of every new
waking type should unavoidably and immediately
cause the apparition of a new type of sleep.
This reasoning could scem to be gratuitous and
unsupported, but it has probably worked more
than once. The telencephaIon has changed two
times in the history of the vertebrate brain,
not only in mammals, but also in birds. We do
not know enough about these animals, however,
for whatever unknown reasons, they followed an
evolutionary path independent from, but parallel
to the mammalian one. After developing a new
telencephalic striate complex of capacity analog
to the mammalian neocortex, they should also
have transformed their old reptilian waking in
SWS, ending with the same two types of sleep:
the appearance of a new type of waking forced,
only by itself, the appearance of again a new
type of sleep.
Considering the final result of why we
sleep, the paradigm defended in this paper
affirms that the particular features of the
mammalian sleep confer (in principle) no
particular advantages over those found in
animals just having activity and rest periods.
The success of new paradignis depends perhaps
only in part on the empirical facts supporting
them and mainly on being able in enduring the
resistance opposed by the defenders of
différent ideas. One should perhaps
consider the possibility of unjustified bias and
preconceptions to maintain time and resource
consuming efforts with extremely low results.
This is undoubtedly, the actual outcome of more
than 50 research years pursuing the answer to
the question on why we sleep. Lets us admit that
science knows for sure that many traits of live
beings are neutral and without adaptive value,
and that the mammalian sleep could just bc
another such example. The most parsimonious
explanation is that the universality of the
activity and rest cycles must have a cause of
higher importance than the existence of a
complex mixture of electrophysiological features
in a particular branch of the animal kingdom. In
any case, the burden of the proof should not be
demanded to those assuming a continuity in the
causes of the sleep, but tothose searching for
additional ones.