One of the more curious aspects observed in
human behavior is the tendency to adopt the
gestures and pos- tures of others (e.g., the
chameleon effect; Chartrand & Bargh, 1999).
Perhaps the most well-known example of this is
contagious yawning (i.e., the act of yawning or
the urge to yawn in the presence of another
person yawning; Provine, 1986). The widespread
familiarity of contagious yawning is due in part
to the fact that almost everyone has experienced
the phenomenon and that it occurs seem- ingly
without volition. Indeed, even the mere thought
of yawning (e.g., reading about it) can be
enough to incite contagious yawning (Provine,
1986). Despite the near uni- versality of this
phenomenon, however, research inves- tigating
the cortical mechanisms underlying contagious
yawning, although providing many advances, has
yet to fully explain its neural basis (Provine,
2005).
A thorough understanding of the neural
underpinnings of contagious yawning is important
for an understanding of the behavioral and
perhaps the communication systems of human and
non human species, and it may be particularly
relevant to revealing why certain human
populations demonstrate absent or aberrant forms
of mimicry (e.g., autism; see Rogers, Hepburn,
Stackhouse, & Wehner, 2003). Recently, a
potential advance in the understanding of
contagious yawning has arisen with the discovery
of mir- ror neurons (MNs). These cells, located
in the posterior inferior frontal gyrus (pIFG)
of monkeys and assumedly humans (Fadiga,
Fogassi, Pavesi, & Rizzolatti, 1995; Riz-
zolatti & Craighero, 2004), become active
when an animal performs a particular action as
well as when it perceives another perform that
same action (di Pellegrino, Fadiga, Fogassi,
Gallese, & Rizzolatti, 1992; Gallese,
Fadiga, Fo- gassi, & Rizzolatti, 1996;
Rizzolatti & Craighero, 2004; Rizzolatti,
Fadiga, Gallese, & Fogassi, 1996).
Accordingly, MNs provide a basis for action
understanding (Rizzolatti & Craighero, 2004)
and may form part of the neural sub- strate
underlying imitative actions (Jeannerod, 1994;
but see Makuuchi,
2005)—capacities both relevant
to the phenomenon of contagious yawning. There
is also speculation that the MN system extends
into visceromotor brain cen- ters, thereby
enabling the experiential understanding of
others, emotional states (Gallese, Keysers,
& Rizzolatti, 2004; Keysers & Gazzola,
2006; Wicker et al., 2003). Here too, a putative
link with contagious yawning exists, because
con- tagious yawning appears to be part of the
neural network involved in empathy (Platek,
Critton, Myers, & Gallup, 2003; Platek,
Mohamed, & Gallup, 2005). Accordingly, there
is reason to speculate that IFG cells may
contribute to the phenomenon of contagious
yawning (Provine, 2005). That said, null
findings from two fMRI investigations of
contagious yawning (Platek et al., 2005;
Schiirmann et al., 2005) have called this
hypothesis into question, leading to speculation
that cells in the pIFG seem to be circumvented
when a yawn is observed. It is possible,
however, that contagious yawn-related
hemodynamic activity in those studies was
obscured by issues of experimental design (e.g.,
block designs and task requirements) that
rendered it impossible to selectively average
only those yawn stimuli that actually elicited
an urge to yawn in each participant. In
addition, it is arguable that some of the
control stimuli used in those experiments were
also MN stimulating, further reducing the
likelihood of observing pIFG activity in
response to observing yawns.
In the present auditory fMRI study, we
reexamined the neural underpinnings of
contagious yawning, this time using an
event-related experimental design in which
participants rated how much each stimulus made
them feel like yawning. In this way, it was
possible to isolate hemodynamic activity
associated with those yawn stimuli that elicited
strong urges to yawn (i.e., contagious yawning)
within each participant. An auditory approach
was adopted primarily because a formal
investigation was needed to support the widely
held, although largely anecdotal (Moore, 1942),
notion that the sound of a yawn elicits
contagious yawning. Like visual images, sounds
are also effective MN stimulators (Gazzola,
Aziz-Zadeh, & Keysers, 2006; Keysers et al.,
2003; Kohier et al., 2002; Pazzaglia,
Pizzamiglio, Pes, & Aglioti, 2008;
Rizzolatti & Craighero, 2004), and so an
auditory paradigm was considered to be a valid
test of the MN hypothesis.
DISCUSSION
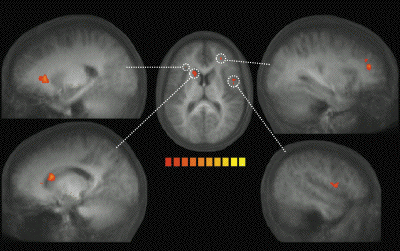
Our results clearly show that hearing
someone yawn not only increases a person's urge
to yawn, but that it activates brain areas that
(1) have been shown to be involved in hearing
and executing mouth actions (Ga.zzola et al.,
2006), and (2) are necessary for recognizing the
actions of others (Pazzaglia et al., 2008).
Accordingly, our findings link contagious
yawning to the human MN network (Rizzolatti
& Craighero, 2004). In fact, the more
contagious the yawn stimulus was, the more these
regions (i.e., pIFG and pSTG) were activated.
Such activity cannot easily be explained by
appealing to notions of yawn suppression (e.g.,
the activation was the result of participants
inhibiting the urge to yawn while undergoing a
movement-sensitive fMRI scanning procedure),
because the yawn stimuli associated with high
urge-to-yawn ratings still generated more BOLD
activity in these regions than did the control
stimuli associated with equivalently high
urge-to-yawn ratings.
These findings are all the more significant
given that prior fMRI studies argued against
pIFG involvement in contagious yawning (Platek
et al., 2005; Schtirrnann et al., 2005). As in
our study, yawh-specific activation had been
observed in the right posterior superior
temporal lobe, perhaps related to the processing
of socially relevant biological motion and its
imitation (lacoboni, 2005; Schiirmann et al.,
2005) or to the fact that the stimulus was a
mouth action sound (Gazzola et al., 2006). But
neither of the previous contagious yawning
studies found that watching people yawn resulted
in differential pIFG activation when compared
with watching people laugh (Platek et al., 2005)
or move their tongue in their cheek
(Schürmann et al., 2005). This has prompted
speculation that contagious yawning is an
automatically released behavior that circumvents
the essential parts of the MN system
(Schürmann et al., 2005).
On reflection, however, it seems possible
that yawnrelated pIFG activity in those studies
could have been masked by activity elicited by
the control conditions. Laughing, for instance,
is also a contagious behavior (Provine, 2005)
that can invoke right pIFG activity, during both
its perception and expression (Meyer, Baumann,
Wildgruber, & Alter, 2007). Similarly, the
tongue-in-cheek control condition used by
Schurmann et al. (2005) may also have elicited
an imitation neural response, given that human
MNs are believed to respond even to movements
that are not goal directed (Rizzolatti &
Craighero, 2004). Indeed, when the results from
that study are reexamined, pIFG activation is
noticeably present for both the yawn and tongue
stimuli (Schürmann et al., 2005, Figure 2,
p. 1262). Moreover, the activation appears to be
stronger in the right IFG and also appears
greatest for the yawn stimuli. In the present
study, the control stimuli (i.e., the scrambled
yawn sounds) may be considered more "action
benign" in the sense that they were artificially
created, physically impossible stimuli.
Second, it is very likely that the block
design nature of the previous studies reduced
the ability to isolate contagious yawn
hemodynamic activity. Clearly, contagious
yawning does not always occur every time a yawn
stimulus has been perceived. Although some yawn
stimuli are more effective than others, a
stimulus that is effective for one person may
not be so for another. Furthermore, there is
most likely variation even within a given
individual from one occasion to the next. Unlike
the previous designs, the online rating
technique of the present eventrelated experiment
enabled us to identify and isolate brain
responses to yawn sounds that were experienced
as contagious compared with those that were not.
That said, it is unclear to what extent the
yawning judgments themselves (i.e., the
urge-to-yawn ratings) affected the activity
observed in response to contagious yawns.
Although we do not deny that being cognizant
of yawning may prime a person to be more
susceptible to contagious yawning, this should
not invalidate the present results. Indeed,
people are known to be more susceptible to
yawning under different circumstances (Provine
& Hamernik, 1986), and many would agree that
there are occasions in which one is more
susceptible to contagious yawning than others
(e.g., in situations were attention is not
strongly focused on a task at hand). This makes
the point that, in some respects, contagious
yawning may rely on some form of embodiment or
mental reflection of the yawn that has been
perceived, be it at its most basic, perhaps
subconscious, level (see also Walusinski,
2006).
Contagious Yawning and MNs
Our finding of contagious yawn-related
activity in what is thought to be the hub of the
MN system (i.e., the pIFG; Rizzolatti &
Craighero, 2004) is not the only datum
consistent with an MN hypothesis of contagious
yawning. First, it has been argued that the
human MN system is not functional at birth but
develops some time after 6 months of age
(Falck-Ytter, Gredebäck, & von Hofsten,
2006). It is noteworthy that, as an antecedent
to any MN-mediated behavior, human contagious
yawning is reportedly not exhibited until some
time after the second year of life (Anderson
& Meno, 2003; Piaget, 1951), despite the
fact that spontaneous yawning is readily
exhibited by infants, newborns, and even fetuses
(Egerman & Emerson, 1996; Sepulveda &
Mangiamarchi, 1995). This incidentally
implicates a perceptual and/or cognitive
component as being central to contagious yawning
(Lejimann, 1979; Platek et al., 2005).
Also, children with autism, a disorder that
may be related to MN-system abnormalities
(Oberman & Ra machandran, 2007), show a
marked reduction in contagious yawning despite
exhibiting "normal" spontaneous yawning behavior
(Senju et al., 2007). Our findings show a
positive correlation between a person's
susceptibility to contagious yawning and how
empathic they are. Furthermore, individuals who
were more empathic showed greater right pIFG
activations to contagious yawn sounds,
consistent with the idea that the MN system is
central to social cognition (Gazzola et al.,
2006; Kaplan & lacoboni, 2006; Pfeifer et
al., 2008). Lastly, akin to the fact that MNs
are suspected only in certain populations (e.g.,
primates; Rizzolatti & Craighero, 2004), it
is interesting that contagious yawning has been
documented in only humans,
chimpanzees (Anderson, Myowa-Yamakoshi,
& Matsuzawa, 2004), and (questionably)
macaques (Paukner & Anderson, 2006),'
despite the fact that a vast array of vertebrate
species are known to exhibit spontaneous yawning
(Baenninger, 1987).
Given that classically defined MN-mediated
behaviors are considered to be those that invoke
mirror cell activity during the observation as
well as the execution of the relevant behavior
(Rizzolatti & Craighero, 2004), one might
expect the pIFG also to be active when a person
yawns. However, because of the stereotypical
head movements associated with the act of
yawning (Deputte, 1994; Provine, 1986; Redican,
1975), this question is difficult to directly
assess with functional neuroimaging.
Nevertheless, there is good reason to believe
that the pIFG may be associated with yawning.
First, it has been reported that a patient with
Foix-Chavany-Marie syndrome, a form of
pseudobulbar palsy resulting from bilateral
opercular lesions, was unable to yawn
voluntarily (whether spared involuntary yawning
was spontaneous and/or contagious is unclear;
Laurent-Vannier, Fadda, Laigle, Dusser, &
Leroy-Malherbe, 1999). Second, and more
convincingly, recent fMRI investigations of
mouth movements have shown that very proximal,
if not the same, regions of the pIFG in BA44 are
activated when participants are asked to smile
(Warren et al., 2006), imitate facial
expressions (Lee, Josephs, Dolan, &
Critchley, 2006; van der Gang, Minderaa, &
Keysers, 2007), manipulate small objects with
their lips (Gazzola et al., 2006), listen to
pure mouth sounds (e.g., crunching a piece of
candy with the teeth, kissing, gurgling,
crunching potato chips, finishing a can of soft
drink with a straw; Gazzola et al., 2006), or
listen to nonverbal human vocalizations (Warren
et al., 2006).
Although what functional or evolutionary
advantage that contagious yawning confers is
unknown, one popular hypothesis is that it
serves as some type of communicative
gesture-that is, to synchronize moods in
gregarious animals (Deputte, 1994), or to
increase vigilance (Daquin, Micallef, &
Blin, 2001; Gallup & Gallup, 2007).
Certainly, the left pIFG is important for the
production of verbal communication signals
(Pa.zzaglia et al., 2008), and the homologous
region in the right hemisphere appears to be
important for nonverbal acoustic signals (Meyer
& Jäncke, 2006). In fact, both brain
regions activated during the perception of
contagious yawns in the present study (i.e.,
right pIFG and pSTG) are thought to be part of a
gesture-recognition network (Villarreal et al.,
2008). In this regard, the right pSTG activation
that correlated with yawn contagiousness ratings
could also be viewed as a node for abstract
(i.e., nonlexical) language comprehension
(Bookheimer, 2002; Gemsbacher & Kaschak,
2003; Jung-Beeman, 2005).
pIFG Neurons: Triggers for Contagious
Behavior?
Inferior prefrontal hemodynamic activity
previously has been shown to increase with the
amount of explicit imitation that a person
exerts (e.g., using a facial imitation task; Lee
et al., 2006), and more recently, this region
has been shown to become active when unconscious
word primes representing actions are presented
to an observer (Galati et al., 2008).
Furthermore, Warren et al. (2006) not only
reported increased pIFG activity when people
passively listened to nonverbal human
vocalizations, they also found concurrent
increases in the electromyographic activity of
lower facial muscles, suggesting that orofacial
gestural responses are engaged automatically
when such stimuli are perceived. In this
context, our novel finding of ventral prefrontal
activity concurrently increasing with contagious
impulses (i.e., the urge to carry out the
behavior) leads to speculation that pIFG
activation may play a role in eliciting the
actual contagious event. Such conjecture will,
of course, require further research.
Finally, as alluded to earlier, it is likely
that attention also has a role in modulating
contagious behaviors. For example, Lee et al.
(2006) found that the inferior prefrontal
activity associated with the imitation of facial
expressions was essentially eliminated when
participants were asked to perform a gender
discrimination task on the same facial stimuli.
This, combined with similar findings from other
studies, prompted Lee et al. to speculate that
the MN system may be attenuated when attention
is focused on anything other than the posture or
behavior to be imitated. Although this too will
require further investigation, one can certainly
imagine a need for such a mechanism. Indeed,
without a way of modulating urges to imitate,
goaldirected activities would be subject to
constant disruption in environments ripe with
social cues.