Jackson SR, Parkinson A, Kim SY,
Schüermann M, Eickhoff SB
.
Department of Brain and
Cognitive Engineering, Korea University,
Seoul
Abstract
Several common neuropsychiatric disorders
(e.g., obsessive-compulsive disorder, Tourette
syndrome (TS), autistic spectrum disorder) are
associated with unpleasant bodily sensations
that are perceived as an urge for action.
Similarly, many of our everyday behaviors are
also characterized by bodily sensations that we
experience as urges for action. Where do these
urges originate? In this paper, we consider the
nature and the functional anatomy of
"urges-for-action," both in the context of
everyday behaviors such as yawning, swallowing,
and micturition, and in relation to clinical
disorders in which the urge-for-action is
considered pathological and substantially
interferes with activities of daily living
(e.g., TS). We review previous frameworks for
thinking about behavioral urges and demonstrate
that there is considerable overlap between the
functional anatomy of urges associated with
everyday behaviors such as swallowing, yawning,
and micturition, and those urges associated with
the generation of tics in TS. Specifically, we
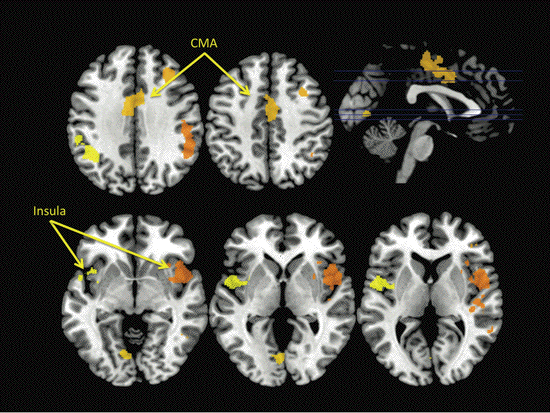
show that the limbic sensory and motor
regions-insula and mid-cingulate cortex-are
common to all of these behaviors, and we argue
that this "motivation-for-action" network should
be considered distinct from an "intentional
action" network, associated with regions of
premotor and parietal cortex, which may be
responsible for the perception of "willed
intention" during the execution of goal-directed
actions.
Many of our everyday behaviors are
characterized by bodily sensations that we
experience either as an urge or a desire for
action. For instance, we may experience a
sensation that our bladder is full that is
accompanied, to a greater or lesser extent, by
an urge or desire to urinate (micturate). In
extreme cases, this sense of fullness can be
quite uncomfortable and the urge to urinate can
be hard to suppress. Similarly, we may
experience a tickle in our throat that is
associated with an urge to cough or to swallow
that can also be difficult to suppress
voluntarily.
However, not all urges for action are
necessarily preceded by bodily sensations of
which we are aware. For example, we may suddenly
experience a strong urge to yawn, or even fmd
ourselves yawning, without being aware of a
sensory "trigger" for the action. In this paper,
we consider the nature and the functional
anatomy of these "urges-for-action," both in the
context of everyday behaviors such as yawning,
swallowing, and urinating, and in relation to
clinical disorders in which the urgefor-action
is considered pathological and substantially
interferes with activities of daily living
(e.g., Tourette syndrome, obsessive-compulsive
disorder, addiction).
A CONCEPTUAL MODEL OF THE
URGE-FOR-ACTION
In common usage, the terms "urge" and
"desire" are frequently encountered as both a
verb (as in to "urge someone on"-i.e., to
motivate, impel, or stimulate a person toward an
action) and as a noun (as in "he felt an urge to
shout"-i.e., a force, drive, or impulse that
impels toward a goal). Furthermore, these terms
are often used interchangeably with one another,
and are frequently listed as synonyms.
Nevertheless, it has been suggested that
there is an important distinction to be made
between urges and desires (e.g., Cameron, 2002;
Davenport, 2008). Thus, Cameron (2002), when
discussing interoception, and the relationship
between conscious awareness and visceral events,
makes a distinction between "detection," which
is an organism's reflexive response based solely
upon afferent physiological information, and
"perception," which refers to an organism's
response based upon all information available to
the organism (which might include learned
information and expectations that might be
generated as a result of learning). Similarly,
Davenport, when discussing mechanisms associated
with the urge-tocough, defines an urge as a
physical need to respond to a sensory stimulus,
and a desire as the translation of an urge into
what he refers to as "a central neural targeted
goal" (Davenport, Sapienza, & BoIser, 2002).
Davenport's motivation-to-action model is
presented in Figure 1.
Combining these two ideas we might conclude
that an urge-as in a drive for action-need not
enter conscious awareness, but that we are
always aware of our desires. An example might
help make this distinction clearer. Imagine that
you are driving your car and you suddenly become
aware of an uncomfortable sensation that your
bladder is full, which you experience as the
urge to urinate. Given that you can in all
likelihood control this urge, you may then form
a desire to urinate in which you construct a
goal or plan which includes a representation of
the behaviors required to complete the action
(e.g., remembering that you passed a filling
station a mile back and knowing that filling
stations usually have public toilets) and a
representation of the likely outcomes of the
action. Can an urge exist if we are not aware of
it? What factors determine whether an urge
enters awareness? How is an urge to act
different from an intention to act? These issues
are discussed below.
ARE URGES SIMPLY REFLEXES?
As outlined above, urges are often defmed as
the drives or impulses that impel us to act.
Nevertheless, it is argued that actions can, and
frequently do, occur in the absence of any
awareness of such drivers, as in the case where
one fmds oneself yawning without previously
being aware of either the desire to yawn or of
any bodily sensation that might reasonably be
identified as giving rise to the yawn. Instead,
one simply fmds oneself yawning. In this case,
it might be argued that this is a reflexive
behavior and not an urge-for-action.
One possible distinguishing feature of
urges, as distinct from reflexes, may be that
urges are chiefly associated with actions that
cannot be realized immediately and must be held
in check until an appropriate time when they
might be released. For instance, when we become
aware of having a full bladder, we experience an
"urgeto-void" because we do not simply void our
bladder, but instead employ a coordinated set of
central, autonomic, and peripheral neural
mechanisms to withhold micturition until we are
in an appropriate behavioral context. Similarly,
in the case of yawning, we might defme the
urge-to-yawn as arising in circumstances where
we are forced to try to stifle the yawn rather
than in the situation where we fmd that we are
yawning.
HOW ARE URGES RELATED TO
AWARENESS?
One factor that may determine whether an
urge enters awareness is the intensity of the
physiological afferent. It has been demonstrated
in the context of the urge-tocough that the
perceived strength of the urge that is
experienced is related to the intensity of
stimulation. Specifically, when capsaicin is
added to the breathing circuit, it results in a
sensation that is perceived as an urge-to-cough.
Furthermore, increasing capsaicin levels leads
to a reliable increase in estimates of this urge
(Davenport, Sapienza, & BoIser, 2002).
Similarly, in our own unpublished studies of the
effects of oropharyngeal stimulation on the
urge-to-swallow, we have found that
oropharyngeal stimulation using pulses of air
produces both an urge-to-swallow and overt
swallowing (for similar findings, see also
Lowell et al., 2008). More importantly, we have
found that increasing the intensity of
oropharyngeal stimulation leads to an increase
in the strength of the perceived
urge-to-swallow. An important question therefore
concerns the role of awareness, and by
implication "desire," in the initiation of
urge-related actions.
According to the motivation-to-action model
represented in Figure 1, actions are initiated
only after a desire to perform an action has
been formed. However, we do not see this step as
necessary and would amend this model, so as to
allow for actions to be initiated directly,
without the need to first construct an explicit
"desire for action." Two factors motivate this
amendment. First, it strikes us that in many
instances actions (e.g., yawning) can be
triggered without our necessarily being aware of
any explicit desire. Second, in many clinical
conditions (e.g., Tourette syndrome), as we
shall see below, unwanted actions or behaviors,
which individuals struggle actively to suppress,
are nevertheless preceded by strong
premonitionary urges. In our view, it is
difficult to reconcile the concept of a
"desire," which is often defmed as "a longing or
craving for something that brings satisfaction
or enjoyment," with unwanted actions, the
execution of which is experienced as unpleasant
and distressing.
If we accept the premise outlined above that
urges-foraction are often accompanied by bodily
sensations, then it strikes us that an important
distinction can be drawn between being aware of
a bodily sensation and being aware of an
urge-for-action. This distinction can be best
illustrated by considering the following
examples. In the case of an itch, we may be
aware of experiencing both an itch (bodily
sensation) and an urge to scratch the itch
(urge-for-action). By contrast, while we may
become aware of an urge-to-yawn, it is not
entirely clear that we are ever aware of the
bodily sensation that gives rise to the urge to
yawn. Furthermore, this becomes more important
if we accept the argument outlined above that
urges occur primarily in circumstances in which
actions may need to be suppressed or their
execution deferred. In such circumstances, we
might distinguish between suppression of
the
action associated with the urge-for-action,
or suppression of the bodily sensation giving
rise to the urge. This distinction has important
clinical implications (see below), and an
interesting issue for future research will be to
detemiine whether the suppression of bodily
sensations and the suppression of
urges-for-action differ in terms of their
functional anatomy.
HOW IS AN URGE TO ACT DIFFERENT FROM AN
INTENTION TO ACT?
The kinds of actions that we have considered
as representative of urges-for-action are highly
automatic, habitual responses that occur
primarily in response to sensory stimulation.
These might include brushing an insect off your
aim, scratching an itch, yawning when tired,
coughing in response to a tickle in your throat,
etc. While such actions can, in some
circumstances, be executed with little or no
awareness of the sensory stimulation that
triggered the action, as when one finds oneself
yawning or coughing, we have argued that a key
characteristic of urges-for-action is that they
involve the suppression or deferment of an
action. Such actions might therefore be
contrasted to intentional, goal-directed, forms
of action.
The circumstances in which the "willed
intention" to execute an action can be shown to
follow the brain processes involved in the
preparation for action were famously studied by
Benjamin Libet (Libet, Gleason, Wright, &
Pearl, 1983), and more recently by Patrick
Haggard and colleagues (e.g., Haggard, 2005;
Haggard & Eimer, 1999; Sirigu et al., 2004).
In Libet's task, participants fixated on a
time-varying, rotating visual spot and were
instructed to make a voluntary hand movement
whenever they felt the "urge" to do so.
Participants were asked to indicate the location
occupied by the moving spot when they had first
felt the urge to move their hand. Libet showed
that this "W judgment" occurred some 200 ms
prior to movement onset, but, more importantly,
he also showed that the preparatory brain
activity that precedes voluntary action, the
socalled "readiness potential," itself preceded
the "Wjudgment" by several hundred milliseconds.
Such readiness potentials arise in the premotor
regions of cortex, including both the
supplementary motor area (SMA) and the
presupplementary motor area (pre-SMA), regions
that have been linked to the planning and
preparation of intentional, goal-directed,
actions and sequences of actions (Deecke &
Kornhuber, 1978).
Haggard has argued that conscious awareness
of our intention to act arises during the
preparatory processes that precede an action,
and is linked to the joint activity of premotor
and parietal brain areas (Haggard, 2005).In
support of this view, he has shown that patients
with damage to the parietal cortex show a
specific impairment in reporting when they
became aware of their intention to move (i.e.,
Libet's "W judgment"). The proposal that the
parietal cortex may maintain a dynamically
updated state estimate of the current postural
configuration of the body (the body schema) is
well supported by neuropsychological (e.g.,
Wolpert, Goodbody, & Husain, 1998) and
recent fMRI studies (e.g., Parkinson, Condon,
& Jackson, 2010; Pellijeff Bonilha, Morgan,
McKenzie, & Jackson, 2006).
While it is clearly the case that in such
experiments individuals can report when they
first perceived themselves to have fonned an
"intention" to move, it is another thing
entirely to argue that such conscious
"intentions" typically precede everyday actions.
Thus, when I am sitting at my desk typing and I
break off to reach for my coffee cup I am not
aware of foiming an intention prior to each
keystroke that I make or of forming an intention
to reach for my coffee. Instead, I am aware of
the actions I am making. Similarly, while it is
clearly demonstrated that our actions are
preceded by neural activity, as discussed by
Libet and by Haggard and colleagues, it is
currently unclear how these activations relate
to the phenomenology of intention.
Haggard and others (e.g., Blakemore,
Wolpert, & Frith, 2002; Haggard, 2005) have
argued that the sense-of-agency that typically
accompanies the execution of voluntary movements
arises as a result of internal forward sensory
models that generate a prediction of the sensory
consequences of an action that is then matched
against afferent sensory signals. It has been
proposed that in cases where the link between
these sensory predictions and confirmatory
sensory input is broken, neurological syndromes
such as anosagnosia (lack of awareness of
injury) or somatoparaphrenia (denial of limb
ownership) may occur (Tsakiris, 2010).
Interestingly, both of these disorders have been
linked to damage of the anterior insular cortex
of the right hemisphere (Baier & Karnath,
2008; Karnath, Baier, & Nagele, 2005).
However, it is important to note that this sense
of agency may in fact have a significant
postdictive or reconstructive component (Moore,
Lagnado, Deal, & Haggard, 2009; Wegner,
2002) and thus is not necessarily an unambiguous
index of intentionality.