Yawning,
acute stressors, and arousal reduction in Nazca
booby adults and nestlings
Amy C. Liang, Jacquelyn K. Grace,
Emily M. Tompkins, David J. Anderson
Department of Biology, Wake
Forest University, Winston-Salem USA
Yawning is a familiar and phylogenetically
widespread phenomenon, but no consensus exists
regarding its functional significance.
The authors tested the hypothesis that
yawning communicates to others a transition from
a state of physiological and/or psychological
arousal (for example, due to action of a
stressor) to a more relaxed state. This arousal
reduction hypothesis predicts little yawning
during arousal and more yawning (above baseline)
during and after down-regulation of
arousal.
Experimental capture-restraint tests with
wild adult Nazca boobies (Sula granti), a
seabird, increased yawning frequency after
release from restraint, but yawning was almost
absent during tests. Natural maltreatment by
non-parental adults also increased yawning by
nestlings, but only after the maltreatment ended
and the adult left.
CORT (corticosterone) was a logical a priori
element of the stress response affecting the
stressor&endash;yawning relationship under the
arousal reduction hypothesis, and cannot be
excluded as such for adults in capture-restraint
tests but is apparently unimportant for
nestlings being maltreated by adults.
The arousal reduction hypothesis unites
formerly disparate results on yawning: its
socially contagious nature in some taxa, its
clear pharmacological connection to the stress
response, and its temporal linkage to
transitions in arousal between consciousness and
sleep.
Gallup
AC, Clark AB Commentary: yawning, acute
stressors, and arousal reduction in Nazca booby
adults and nestlings. Fontiers in Psychology 29
october 2015
Les auteurs ont testé
l'hypothèse que le bâillement
communique aux autres une transition d'un
état d'excitation physiologique et / ou
psychologique (par exemple, grâce à
l'action d'un facteur de stress) à un
état plus détendu. Cette
hypothèse de réduction de
l'excitation prédit peu bâillements
pendant l'éveil et plus bâillements
pendant et après la régulation de
l'excitation.
Les expériences ont été
menées avec des oiseaux de mer adultes
sauvages "les fous de Nazca (Sula de granti)".
D'abord saisis, puis maintenus en cage, c'est
après leur libération que
l'augmentation de la fréquence des
bâillements a été
mesurée. Lors des test initiaux, les
bâillements étaient presque
inexistants.
Après avoir constaté que dans
la nature, des oisillons subissent de mauvais
traitements de la part d'adultes autres que
leurs parents, les bâillements des
oisillons ont été
comptabilisés dans ces circonstances.
L'augmentation du nombre de bâillements
intervient après la fin de la
maltraitance quand l'adulte responsable est
parti.
La corticostérone, un des facteurs de
la réponse au stress, affecte a priori la
relation stress - bâillements dans
l'hypothèse de l'effet apaisant
post-stress du bâillement. Ceci est apparu
probable chez les adultes après capture
mais n'a pas été confirmé
pour les oissilons victimes de
maltraitance.
L'hypothèse de réduction de
l'état de stress permet de rassembler des
résultats autrefois disparates concernant
le bâillement: sa nature socialement
contagieuse dans certains taxons, son lien
pharmacologiquement avéré avec la
réponse au stress, et son lien temporel
des états transitionnels entre
l'éveil et le sommeil.
1. Introduction
Although yawning is ubiquitous in humans and
phylogenetically widespread, studies targeting
functional significance have produced little
consensus. The wide variety of functional
explanations includes increasing alertness,
renewing surfactant in the lungs, and evacuating
infectious substances fromthe tonsils. The
popular viewthat yawning adjusts oxygen and
carbon dioxide levels in the blood persists,
despite the demonstration by Provine et al. that
human subjects breathing an elevated level of
oxygen did not yawn more or less than did
subjects breathing air with elevated carbon
dioxide. Our goal in this paper is not to review
this extensive and diverse literature;
instead,we focus on one idea, that yawning and
recent exposure to a stressor are linked (by
"stressor" we mean an extrinsic factor, real or
perceived, that threatens homeostasis). A
variety of evidence connects yawning, or at
least behavior that resembles yawning, to
stressors, both physiological and social.
The HPA
(Hypothalamus&endash;Pituitary&endash;Adrenal)
axis response via glucocorticoids, a widely
investigated aspect of the stress response, is
of special interest. Adrenalectomy stops
production of glucocorticoids and virtually
abolishes yawning in rats; administration of the
synthetic glucocorticoid dexamethasone to these
same rats restores yawning. The hormones
dopamine and oxytocin are both products and
regulators of the HPA axis; exogenous
administration of these hormones or their
agonists induces yawning. Circulating levels of
corticosterone (CORT), the primary
glucocorticoid in birds, and cortisol (the
mammalian homologue of CORT) are also correlated
with alertness, vigilance, and arousal, rising
in the hours before awakening, rising faster
just after awakening, and promoting arousal and
memory consolidation.
Dourish and Cooper linked yawning and both
of these actions of the HPA axis, suggesting
that yawning "may signal the termination of
stressful experience or of sustained
concentration", an idea that we call the
"arousal reduction hypothesis" to distinguish it
from Baenninger's suggestion that yawning
increases arousal. Under this hypothesis,
adaptive down-regulation of arousal also causes
yawning and occurs when an animal perceives that
a stressor has passed and that arousal in
response to the stressor is no longer required.
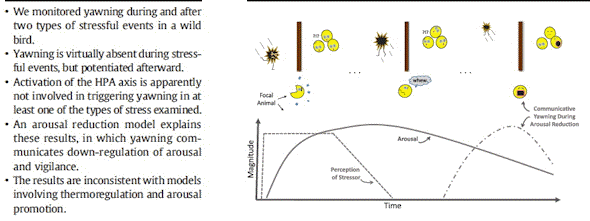
Here we test the arousal reduction hypothesis,
measuring yawning in response to temporary
stressors in a wild bird, the Nazca booby (Sula
granti), using both experimental and natural
acute stressors. Wild Nazca boobies tolerate
close human presence and handling well,
facilitating behavioral observation even after
experimental administration of stressors.
In adults, a standardized capture-restraint
stress test induced an HPA axis response,
raising the circulating concentration of CORT
.We monitored the birds' behavior, including
yawning, during the test and after release from
restraint and compared the yawning rate to that
of undisturbed adults. In nestlings, we
exploited the natural phenomenon of episodes of
nestling maltreatment by Non-parental Adult
Visitors (NAVs). NAVs exhibit an intense social
attraction to nonfamilial conspecific nestlings,
which frequently manifests as aggressive
interactions with nestlings, with a gradation in
severity.
We compared the yawning of nestlings that
experienced a low severity aggressive NAV
interaction, which induces thermoregulatory
stress, to that of time- and age-matched
undisturbed nestlings. We predicted low rates of
yawning during and shortly after the period of
stress and a higher rate after a period of
surveillance following removal of the stressor.
If CORT is the target of the down-regulation
under this hypothesis, then we predicted that
only a stressor that induced elevated
[CORT]would induce subsequent yawning
and that [CORT] during the period of
stress would predict the subsequent
yawning.
4. Discussion
This study is the first to evaluate known
stressors as triggers of yawning in a wild,
free-living animal; the results are consistent
with the arousal reduction hypothesis, while
indicating that the target(s) of arousal
down-regulation could include circulating CORT
but does not in at least some contexts. Adult
Nazca boobies had higher circulating
[CORT] during nocturnal
capture-restraint tests, demonstrating
activation of the HPA axis.
Our Control adults showed that yawning is
rare at night and during capture-restraint
tests, but these adults all yawned at least once
in the 60 min following the capture-restraint
stressor, while time-matched controls did not.
Nestlings were prevented from thermoregulating
in this tropical environment during diurnal
visits from NAVs, at a time of day when solar
insolation induced all time-matched, unvisited
nestlings to dump heat continuously by gular
flutter. The social and thermal stressors of
these low-severity NAV events did not induce a
detectable [CORT] response in nestlings
(in contrast to high-severity NAV events), but
did induce yawning (and thermoregulation by
gular flutter) in all Post-NAV nestlings within
15 min of the NAV's departure . Thus, two types
of stressor reliably triggered yawning, in the
temporal sequence predicted by the arousal
reduction hypothesis, and a detectable CORT
response was not required in one of the
stressor&endash;yawning relationships.
Our results provide insight into the
cause-and-effect relationship of stressors and
yawning. If [CORT] caused the yawning
response in adults then the large variation in
CORT response during the test might predict the
latency to yawn, but no predictive ability was
detected. Instead, the latency to yawn after the
test showed little variation, consistent with a
yes/no trigger by the stressor (which could be
CORT, if its effect is qualitative). The thermal
stressor affecting nestlings also showed
apparent quantitative variation: some nestlings
abandoned the pelican posture and stood upright
immediately after their NAV's departure, while
others waited up to 7 min to stand (Fig. 4),
probably reflecting variation in perception of
the end of the risk from the NAV. All control
nestlings dumped heat via gular flutter for 100%
of the observation period, and all victim
nestlings resumed gular flutter within minutes
of standing, so the variation in latency to
stand probably does not reflect meaningful
variation in thermal load incurred before the
NAV visit, and the added load in pelican posture
during the visit was presumably similar for all
victims. Instead, latency to stand should serve
as a proxy for cumulative thermal load (longer
latency extends the period without gular
flutter), and the latency to the first yawn
after standing showed a significant positive
relationship with the latency to stand. Thus,
the yawning response to a stressor can be
qualitative or quantitative, depending on
context.
Several hypotheses have been proposed to
explain the causation of yawning. Under the
thermoregulation hypothesis, yawning is
triggered by brain hyperthermia and cools at
least part of the brain via targeted
ventilation.Our results from adults are silent
on this issue because we have no data on the
adults' temperatures. For nestlings, this
hypothesis predicts a negative relationship
between latency to stand (a proxy for
hyperthermia) and latency to yawn after
standing, but we observed a significant positive
relationship.
The arousal promotion hypothesis casts
yawning as a means of regulating arousal,
increasing attention to surroundings "when
environments provide relatively little
stimulation"; humans use yawning to counter
boredom under this hypothesis. [CORT]
exhibits correlation with the sleep&endash;wake
cycle (for Nazca boobies,) consistent with a
primary role in arousal/attention. Nazca booby
adults exhibited variation in [CORT] at
the end of their restraint, so this hypothesis
predicts a quantitative, negative relationship
between [CORT] and latency to yawn,
which was not observed. Noting that Post-Test
adults and Victim nestlings in this study
experienced significant stimulation only minutes
before their first yawn, and gave no indications
of drowsiness, the essential assumption of the
arousal promotion hypothesis seems to be largely
invalid in our study.
Dourish and Cooper's arousal reduction
hypothesis is the reverse of the arousal
promotion idea. The results of several studies
support this hypothesis. In these studies the
animal was released from a stressor, had time
for surveillance of the stressor and a decision
that the stressor had passed, and then showed
enhanced yawning. For Nazca booby adults,
yawning was essentially absent during the
recapturerestraint test and for 30 min after
release, and then potentiated in the next 30
min. Nestlings also showed an
inhibition/potentiation cycle: despite probable
hyperthermia, 18 of 27 (0.67) nestlings visited
by NAVs showed a latency to yawn of at least 1
min after assuming a normal posture that would
permit gular flutter. In a study with a similar
design, budgerigars (Melopsittacus undulatus)
given a capturerestraint test suppressed yawning
during the first 20 min after release, then
increased yawning in the next 20 min, and
finally returned to a control level 40&endash;60
min after release. Noting a correlation between
yawning and body temperature in their
experiment,Miller et al. suggested that yawning
has a thermoregulatory function.
The arousal reduction hypothesis offers an
alternate interpretation: after a period of
alert surveillance, the stress response was
down-regulated adaptively and signaled by
yawning when the bird perceived that the
stressor is gone. Finally, rats (Rattus rattus)
experiencing shocks at predictable intervals
(during which down-regulation of the stress
response can be advantageous) yawned more
frequently between shocks than did rats
experiencing shocks at random intervals (during
which down-regulation is not advantageous),
suggesting an association between yawning and
down-regulation of stress. Moyaho and Valencia
concluded that yawning diminished arousal,
exerting a calming effect, but the reversed
arrow of causation posited by the arousal
reduction hypothesis, that diminishing arousal
causes yawning, is equally consistent with their
results.
The arousal reduction hypothesis attributes
communication value to the signal of a
subjective perception that a stressor has
passed: the animal is sending a message actively
about its perception. This position connects
logically to the well-known contagion of yawning
in both non-humans and humans, and to the
connection of susceptibility to contagious
yawning and empathy for the emotion and
motivation of others. As such, the arousal
reduction hypothesis can be considered an
element of the social/communication model for
yawning combined into a single model of adaptive
information-sharing of perceptions of threat
status. Any animal that benefits from
coordination of group members' attention to
surroundings (c.f., even ephemeral groups like
sexual pairs) can be expected to attend to a
signal like yawning and to benefit from sending
the signal in some situations. The more
important such coordination is, the more likely
propagation of the signal (contagion) should be.
The fact that animals yawn when alone is not a
difficulty of this model, possibly indicating
that the generator of yawns lacks a negative
feedback mechanism that checks for the presence
of receivers of the signal.
We suggest a change in the sense of the
hypothesis. Rather than signaling the
"termination of stressful experience or of
sustained concentration", the hypothesis should
focus explicitly on the internal state of the
animal: simply, yawning signals an attempt to
down-regulate arousal. This focus on internal
state, rather than extrinsic factors, admits the
possibilities of adaptive down-regulation of
arousal 1) even when the arousing stimulus still
exists or
2) when no stressor is involved (for
example, at the end of the normal awake period).
A variety of phenomena are apparently consistent
with this hypothesis. Common experience makes us
associate impending sleep with yawning;
downregulating arousal should facilitate the
transition to sleep. Many athletes yawn
immediately before a competition (speed-skater
Apolo Anton Ohno is a notorious example),
perhaps actively calming themselves at a time of
on-going high tension and producing yawns as an
epiphenomenon.
Captive budgerigars yawn more under ambient
temperatures increasing from 27&endash;33
°C than from 23&endash;27 °C or
decreasing from 34&endash;28 °C or
decreasing from 28&endash;24 °C, perhaps
indicating an attempt to suppress agitation in a
situation with almost no options to dump heat.
Yawning contagion propagates differentially from
social leaders to others in highly social
species like bonobos (Pan paniscus), chimpanzees
(P. troglodytes), and ostriches (Struthio
camelus) and is connected to stimulation of
brain areas associated with empathy and social
interpretation in "infected" individuals,
suggesting systematic top-down signaling as part
of yawning's communicative value. In humans,
"evening types" display a relatively nocturnal
circadian cycle and claim difficulty in the
process of awakening and beginning the day, the
opposite of "morning types"; the awakening
stressor for evening types is associated with a
yawning frequency at awakening that is several
times that of "morning types", for whom
awakening is apparently not a stressor,
consistent with the arousal reduction
hypothesis. In conclusion, acute stressors
initially suppressed and later potentiated
yawning in Nazca boobies in a temporal sequence
consistent with elevated arousal followed by
active down-regulation of arousal. CORT was a
logical a priori factor affecting the
stressor&endash;yawning relationship under the
arousal reduction hypothesis, and cannot be
excluded as such for adults in capture-restraint
tests but is apparently unimportant for
nestlings being maltreated by adults. Little
information exists for any species to enable
rigorous tests of the arousal reduction
hypothesis, and we suggest experimental
manipulation of arousal, especially in highly
social species in which coordination of arousal
will be favored by natural selection, and more
careful monitoring of arousal state than was
possible in our study.