The sensitivity of elasmobranchs to weak
electric fields is mediated by small
electroreceptive structures known as the
ampullae of Lorenzini. These sensory vesicles
contact the surrounding water via jelly-filled
canals that lead to groups of pores on the
surface of the head, and enable the animal to
detect voltage gradients as low as 0.01 MV/cm
(Kalmijn, 1966). This sensory modality is used
by sharks for detection or bioelectric fields
produced by prey and may also serve in
electroorientation by geomagnetic induction
(Kalmijn, 1971, 1973).
Previous studies on the electric detection
of prey by sharks were conducted either entirely
in the laboratory or under semi-natural
conditions in the field. In these experiments
animals were first motivated to feed by exposure
to an odor stimulus and then presented
electrical test stimuli (Kalmijn, 1971, 1978).
This report presents the results of preliminary
experiments on predator-prey interactions
between the swell shark, Cephaloscyllium
ventriosum (Garman) (Scyliorhinidae), and the
blacksmith, Chromis Junctip-innis (Cooper)
(Pomacentridae), in their natural habitat, and
supports the postulate that electric fields
produced by small fish mediate successful prey
detection and capture by sharks.
Study area, methods and result.
This study was conducted at a small offshore
island, Ship Rock, located near the Isthmus area
of Santa Catalina Island, California. Here, the
underwater habitat is dominated by a rocky
boulder bottom and large stands of giant kelp,
Macrocystis pyrifera (Linnaeus) C. Agardh.
Blacksmith (approximately 10-15 cm SL) form
large diurnal feeding aggregations in the water
column at the seaward edge of the kelp forest.
At dusk, these assemblages disperse when the
individuals descend to the bottom and refuge
among rocks and benthic macroalgae. During
spawning season in late spring and summer
densities of blacksmith are highest and many
individuals rest on the exposed bottom at night
due to limited shelter.
Swell sharks in shallow waters (<20 m) at
Ship Rock numbered approximately 125
individuals. Sharks rested deep within rock
crevices during the day, and although capable of
active swimming, most individuals rested
motionless on the open bottom at night during
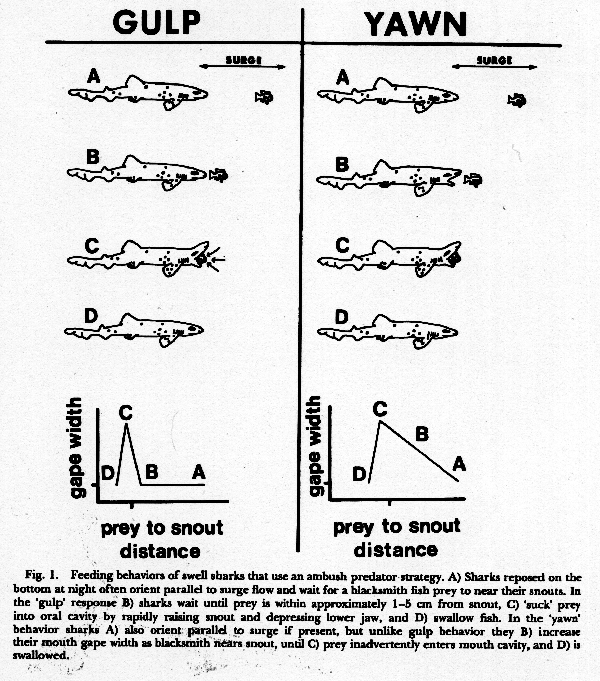
this study (July 1977). Reposed sharks use an
ambush strategy to prey on blacksmith at night,
and capture fish by one of two different feeding
patterns. The first is a rapid 'gulp' behavior
that occurs when a blacksmith comes within l-5
cm of the shark's snout. In this behavior, the
shark rapidly raises its head, depresses its
lower jaw, and sucks the fish into its oral
cavity (Fig. 1). As the mouth closes, water is
expelled through the gill slits and the prey is
swallowed. This feeding mode occurs so rapidly
that only the snap of the shark's mouth can be
seen, not the actual ingestion of the prey. The
second mode of capture is a readily visible
'yawn' behavior that is clearly
discernible from the 'gulp' by a graded
relationship between the shark's gape
width and the prey-to-snout distance. The closer
a blacksmith comes to the head of the shark, the
wider the shark opens its mouth. If the prey
moves away from the snout, the mouth begins to
dose. In the 'yawn' behavior, blacksmith
are captured when they inadvertently swim, or
fortuitously drift in the shallow surge, into
the fully expanded oral cavity of the shark
which is then snapped shut (Fig. 1).
To further investigate these interactions, a
preliminary series of simple underwater tests
were performed to determine how readily sharks
would take free-swimming blacksmith and whether
the sharks used sensory cues other than vision
to detect their prey. The former was tested by
tethering a live healthy blacksmith to the end
of a fiberglass rod with a short piece of
monofilament line. The fish, which did not
struggle, was presented by divers to sharks at
rest on the bottom at night. In this test, seven
out of ten sharks attempted to take the prey.
Next, a live blacksmith was sealed in a
waterfilled dear-plastic bag (that blocked all
olfactory and electrical cues), suspended on the
end of the fiberglass rod, and then offered to
ten sharks. This time, no shark showed any
positive response. Replicate tests using
blacksmith in a black plastic bag produced the
same negative results. These findings indicated
that neither vision (either normal or augmented
by our dive lights) nor olfaction was a primary
sensory modality used to detect prey.
To determine which sense is most important
for detection of blacksmith, a prey chamber was
constructed in which further combinations of
cues could be manipulated. The chamber design
was a modification of that used by Kalmijn
(1971) so that tests similar to his could be
performed. The housing was made from a 5.5-cm
length of 14-cm outside-diameter PVC pipe (.7cm
wall thickness) backed with a removable plastic
plate. The forward end of the housing was strung
with monofilament line to form a webbed face. An
attachable agar cap was made from a 4.7-cm long
14.2-cm inside-diameter PVC pipe and faced on
one end with a covering of 1-cm thick open-cell
foam that had been impregnated with a 3%
agar-seawater gel. The cap, when used, was
placed over the webbed end of the chamber and
sealed by an o-ring seated on the perimeter of
the main prey housing. An additional plastic
shield made of household plastic wrap
approximately 10-p thick was placed over the
face of the agar cap and secured with a flat
elastic band. The entire apparatus was mounted
on the end of a 1-m long plastic pipe for
presentation by divers to sharks.
A live blacksmith was placed in the housing
and presented to sharks encountered by divers
swimming casually around Ship Rock at night. Of
14 sharks presented with an open-ended chamber
containing a healthy fish, 8 (57%) showed a gulp
or yawn response (Table 1). Of ten sharks
presented with the empty (control) chamber (with
and without the agar cap), none showed any
positive feeding response. The permeability of
the agar shield to an electric field was tested
by presenting naive sharks a blacksmith in the
chamber with agar cap affixed. Three of seven
sharks showed a positive feeding response which
indicated the electric fields were penetrating
the agar screen. These experiments showed that
1) feeding responses were due to the presence of
a fish in the chamber and not a reaction to the
apparatus, and 2) not all sharks would show a
positive feeding response, possibly due to a low
state of hunger or an unnatural (e.g., weakened
or distorted) electric field.
During three nights of the following week,
ten sharks chosen at random were tested for
their sensitivity to bioelectric fields of prey.
A shark was first presented with a healthy fish
in an uncovered prey chamber. If the shark
showed a positive response, the agar-faced cap
was placed over the open end of the prey chamber
and again presented to the same shark. With the
cap attached all olfactory and visual cues from
the prey (which did not struggle) were
obliterated; the only stimuli present was the
blacksmith's bioelectric field that passed
through the agar (and possibly some acoustic
cues). Of the ten sharks (out of approximately
25 tested) that showed the initial positive
feeding response, six showed an additional
'gulp' or 'yawn' when presented with the
chamber plus the agar cap (Table 1). After
exposure to the agar-covered chamber, each shark
was once again presented the apparatus with an
additional polyethylene-plastic shield placed
over the agar cap that now screened the electric
field emitted by the prey but still allowed any
acoustic cues present to penetrate. In this
test, no sharks responded to the apparatus.
Table 1 feeding responses of
swell sharks to bioelectric fields
from blacksmith in chamber.
Sharks at rest on the bottom at
night were presented chamber
apparatus by diver. In the prey
electric-field experiments, divers
made serial presentations of the
apparatus (under three different
conditions) to ten individual sharks
so that each shark was given the
same test.
Discussion.
These experiments indicate that detection of
blacksmith by swell sharks using an ambush
strategy under normal nocturnal feeding
conditions can be successfully mediated by the
passive electric field emitted by individual
prey. This sensory modality provides graded
analog information of prey distance at close
ranges (20 cm). This was evident from the
gapesize relationship to prey-to-snout
distance seen in the 'yawn' behavior.
Passive electric field strengths emitted by
small fishes [e.g., 0.2 V/ cm at 10 cm
distance for a 15 cm long flatfish, Pleuronect
sp. (Kalmjn, 1974)], at distances up to 20
cm are within the sensitivity range of small
sharks (.01 AV/cm) (Kalmijn, 1966).
Data from the experiment (Table 1) show a
trend for a decreasing proportion of positive
feeding responses with each successive
presentation. Based on this response decline,
such a trend might be interpreted as a learning
response, i.e., that some sharks had learned
they could not get the fish and therefore no
longer responded. This explanation is inadequate
however, because sharks would have to learn to
associate the apparatus with food in one trial
and then show extinction in the very next
presentation. This is highly unlikely since many
trials (e.g., tens or hundreds) may be necessary
to establish learning in fishes and extinction
may take weeks or months (Harlow, 1939;
O'Connell, 1960; Clark, 1963). It is much more
probable that the response decline was due to
distortion or weakening of the electric field
caused by the agar cap, or perhaps differential
states of motivation among sharks due to prior
feedings, fatigue, etc. In addition, sharks were
observed to be unsuccessful in multiple attempts
to capture blacksmith when feeding naturally,
and no such response declines were
observed.
In contrast to the 'yawn', some
sharks captured prey by the more stereotyped
'gulp' behavior, but only when prey came within
approximately 3 cm or less of the shark's snout.
Although these two predatory modes serve similar
functions, they are qualitatively distinct in
their motor patterns and show different
relationships to prey distance. Further
high-speed cinematographic analysis would
provide more precise data on jaw mechanics and
temporal sequences. It also remains undear
whether the proximate causal factors that
determine which specific feeding behavior is
displayed are environmental (e.g.,
characteristics of electric cue from prey),
intrinsic (e.g., motivational state of the
animal), or a combination of both.
Undoubtedly, predation is frequently
mediated by combinations of different sensory
systems. There are, however, advantages for an
electric sensitivity to prey, especially for a
nocturnal predator. While olfactory and acoustic
cues may be important in initial prey perception
over longer distances, electric field vectors,
like vision, can theoretically provide almost
instantaneous information on direction of prey
at dose range and quantitative information of
distance based on the configuration (or
strength) of the electric field.
References
CLARK, E. 1963. The maintenance of sharks in
captivity, p. 115-149. In. Sharks and survival.
P. W. Gilbert (ed.). D. C. Heath, Boston.
HARLOW, H. F. 1939. Forward conditioning,
backward conditioning, and pseudo-conditioning
in the goldfish. J. Genet. Psychol.
55:49-58.
KALMIJN, A. J. 1966. Electro-perception in
sharks and rays. Nature (Lond.)
212:1232-1233.
KALMIJN, A. J. 1971. The electric sense of
sharks and rays. J. Exp. Biol. 55:371-383.
KALMIJN, A. J. 1973. Electro-orientation in
sharks and rays: theory and experimental
evidence. Scripps Inst. Ocean. Ref. Ser., Contr.
73-39:1-22.
KALMIJN, A. J. 1974. The detection of
electric fields from inanimate and animate
sources other than electric organs, p. 147-200.
In: Handbook of sensory physiology, vol. 11113.
A. Fessard (ed.). Springer-Verlag, Berlin.
KALMIJN, A. J. 1978. Electric and magnetic
sensory world of sharks, skates, and rays, p.
507-528. In: Sensory biology of sharks, skates,
and rays. E. Hodgson and R. Mathewson (eds.).
ONR, U.S. Gov. Printing Office, Wash., DC.
O'CONNEL.L, C. P. 1960. Use of the fish
school for conditioned response experiments.
Anim. Behav. 8: 225-227.