The effect of N-methyl-D-aspartic acid
(NMDA),
(+-)-alpha-amino-3-hydroxy-5-methyl-isoxazole-4-propionic
acid (AMPA), or
(+-)-trans-1-amino-1,3-cyclo-pentanedicarboxylic
acid (ACPD) (5-60 ng in 0.3 microliter of
saline) microinjected in the paraventricular
nucleus of the hypothalamus on penile erection
and yawning was studied in male rats. NMDA
induced both penile erection and yawning in a
dose-dependent manner. AMPA and ACPD also
induced penile erection but less potently than
NMDA, but were ineffective in causing yawning.
NMDA effect on penile erection and yawning was
prevented by (+)-MK-801 (0.05-0.1 mg/kg IP, 10
min before NMDA), by the oxytocin antagonist
d(CH2)5Tyr(Me)-Orn8- vasotocin (50-100 ng ICV 10
min before NMDA), but not by haloperidol
(0.1-0.5 mg/kg IP 10 min before NMDA). The
results suggest that NMDA induces penile
erection and yawning by increasing oxytocinergic
transmission by acting in the paraventricular
nucleus of the hypothalamus.
Penile erection and yawning are two distinct
behavioral patterns that often occur
concomitantly under certain physiological and
experimental conditions. Among substances that
induce both penile erection and yawning,
dopaminergic agonists, oxytocin and
adrenocorticotropin (ACTH) are certainly the
most widely known. While several lines of
experimental evidence suggest that a
dopamine-oxytocin link plays a key role in the
expression of these behavioral responses, ACTH
and related peptides seem to act with a
mechanism(s) not involving central dopamine or
oxytocin.
Accordingly, both penile erection and
yawning are induced by apomorphine, a classical
dopaminergic agonist, and by oxytocin injected
in the paraventricular nucleus of the
hypothalamus (PVN), and both responses are
prevented either by oxytocin antagonists or by
electrolytic lesions of the PVN that deplete
oxytocin in the central nervous system. The
above responses seem to be mediated by an influx
of Ca 2+ ions through voltage-dependent Ca 2+
channels rather than by an increased excitatory
amino acid neurotransmission, because they are
prevented by the intracerebroventricular (ICV)
and PVN injection of w-conotoxin, a potent
inhibitor of voltage-dependent Ca 2+ channels in
the nervous tissue, but not by several types of
excitatory amino acid receptor antagonists.
However, the above results do not rule out the
possibility that excitatory amino acids might
control the expression of the above responses by
acting before apomorphine and/or oxytocin.
Indeed, the PVN receives glutamatergic and/or
aspartatergic innervations from several brain
areas, and recent studies have shown that
excitatory amino acids, i.e., kainic acid and
N-methyl-D-aspartic acid (NMDA) induce yawning
and genital grooming when injected in the PVN.
This prompted us to study the effect of agonists
of the various excitatory amino acid receptor
subtypes when injected in the PVN on penile
erection and yawning in male rats to verify
whether the above behavioral responses to these
excitatory amino acids were mediated by the
activation of dopaminergic and/or oxytocinergic
transmission in the PVN.
Discussion
The present results show that NMDA, a
selective agonist of the excitatory amino acid
NMDA receptor subtype, but not AMPA or ACPD,
agonists of the AMPA and metabotropic receptor
subtypes, respectively, induces penile erection
and yawning when injected in the PVN of male
rats. This is in agreement with previous studies
showing that NMDA and kainic acid induce yawning
and genital grooming when injected in the PVN.
Most important, in agreement with the scarce
effect of either AMPA or ACPD on penile erection
and yawning, NMDA is apparently acting by
stimulating specific NMDA receptors, because it
is prevented by MK-801, a potent noncompetitive
NMDA receptor antagonist, given either IP or in
the PVN. The activation of these NMDA receptors
in the PVN would cause, in turn, the activation
of central oxytocinergic transmission, because
penile erection and yawning induced by NMDA is
prevented by the ICV injection of
d(CHz)sTyr(Me)-OrnS-vasotocin, a potent oxytocin
antagonist. However, NMDA effect would not be
mediated by the release of oxytocin in the PVN,
but in other brain areas, because it is not
prevented by the injection of the oxytocin
antagonist directly in this hypothalamic
nucleus. The possibility that NMDA effect is
mediated by the release of dopamine from
incertohypothalamic neurons impinging on and
activating, in turn, oxytocinergic neurons in
the PVN is also unlikely. In fact, the blockade
of dopaminergic receptors by haloperidol was
found to be unable to prevent NMDAinduced penile
erection and yawning, thus excluding the
existence of a glutamic acid-dopamine-oxytocin
link playing a role in the expression of these
responses. Nevertheless, the results suggest the
existence of a neuronal glutamic acid-oxytocin
link involved in the expression of these
behavioral responses in addition to the already
described dopamine-oxytocin link. It is likely
that the oxytocinergic neurons mediating penile
erection and yawning activated by NMDA are the
same activated by apomorphine and other
dopaminergic agonists, although further studies
are necessary to confirm such a
possibility.
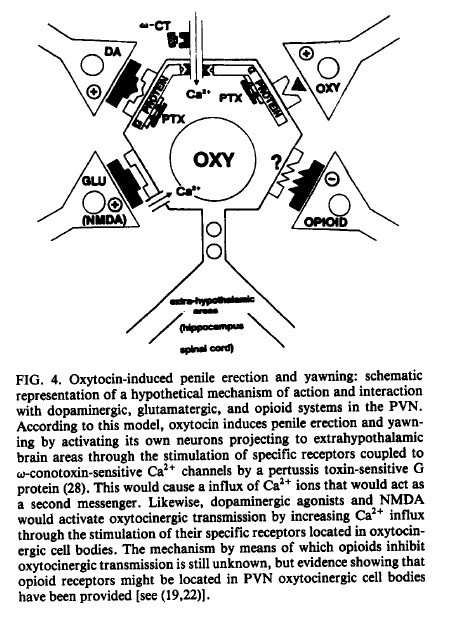
The molecular mechanism by means of which
NMDA activates oxytocinergic transmission is
unknown, and only some speculation is possible
at present. One possibility is that NMDA induces
a Ca 2+ influx through the well-characterized
NMDA receptor-coupled Ca 2+ channel. In this
respect, it is noteworthy that apomorphine and
oxytocin effects also seem to be mediated by a
Ca 2+ influx through o~-conotoxin- sensitive Ca
2÷ channels (3). Interestingly,
NMDA-induced Ca 2+ influx seems to be correlated
with the activation of nitric oxide
(NO)-synthase, the enzyme that produces the
novel discovered neurotransmitter/neuromodulator
NO, and NO-synthase inhibitors prevent
apomorphine- and oxytocininduced penile erection
and yawning. This raises the possibility that
NMDA-, apomorphine-, and oxytocin-induced penile
erection and yawning are mediated by an
increased production of NO secondary to a common
mechanism, e.g., a receptor-induced Ca 2+
influx.
The present study does not support our
previous suggestion that excitatory amino acids
are not involved in the expression of penile
erection and yawning. Our proposal was sustained
by the inability of several excitatory amino
acid receptor antagonists to prevent
apomorphine-, oxytocin-, and ACTHinduced penile
erection and yawning when injected ICV or in the
PVN. The present results, in fact, shows not
only that excitatory amino acids are involved
but also that they activate directly
oxytocinergic transmission to induce these
behavioral responses. Nevertheless, the results
are in line with those of our previous study,
because if one admits that exogenous oxytocin
acts on specific oxytocinergic receptors in the
PVN to induce penile erection and yawning,
excitatory amino acid receptor antagonists would
have not prevented oxytocin effect unless
mediated by the release of an endogenous
excitatory amino acid.
In conclusion, NMDA induces penile erection
and yawning when injected in the PVN. This
suggests that oxytocinergic neurons mediating
these responses are also under the control of
excitatory amino acids in addition to other
neurotransmitters, i.e., dopamine, opioid
peptides, and oxytocin itself, as already
described for PVN oxytocinergic magnocellular
neurons. In particular, dopamine, glutamic acid,
and oxytocin itself seem to have a facilitatory
and opioids an inhibitory role in the expression
of penile erection and yawning.