Studies of contagious yawning have reported
inconsistent findings regarding whether dogs
exhibit this behavior and whether it is mediated
by social-cognitive processes or the result of
physiological arousal. We investigated why some
dogs yawn in response to human yawns;
particularly, whether these dogs are exceptional
in their ability to understand human social cues
or whether they were more physiologically
aroused. Sixty shelter dogs were exposed to
yawning and nonyawning control stimuli
demonstrated by an unfamiliar human. We took
salivary cortisol samples before and after
testing to determine the role of arousal in yawn
contagion. Dogs were tested on the object-choice
task to assess their sensitivity for
interpreting human social cues. We found that 12
dogs yawned only in response to human yawns
(i.e., appeared to exhibit yawn contagion),
though contagious yawning at the population
level was not observed. Dogs that exhibited yawn
contagion did not perform better on the
object-choice task than other dogs, but their
cortisol levels remained elevated after exposure
to human yawning, whereas other dogs had reduced
cortisol levels following yawning stimuli
relative to their baseline levels. We interpret
these findings as showing that human yawning,
when presented in a stressful context, can
further influence arousal in dogs, which then
causes some to yawn. Although the precise
social-cognitive mechanisms that underlie
contagious yawning in dogs are still unclear,
yawning between humans and dogs may involve some
communicative function that is modulated by
context and arousal.
Introduction
Over the last decade, there has been renewed
interest in understanding the mechanisms that
control contagious yawning (i.e., yawning in
response to perceiving another individual yawn).
It has been suggested that yawning functions to
increase arousal in situations in which
environmental stimulation is low and wakefulness
and vigilance are important (Baenninger 1997).
This behavior may have evolved a communicative
mechanism when exhibited within group contexts,
serving to synchronize group activities by
communicating sleepiness or stress (Deputte
1994; Guggisberg et al. 2010). Although it has
been argued that contagious yawning is nothing
more than a behavioral fixed action pattern
(Provine 1986), more recent studies have
indicated that a social-cognitive mechanism
(i.e., empathy) is involved in contagious
yawning. Humans who score higher on empathy
scales (Arnott et al. 2009) and faux pas theory
of mind and self-recognition tasks (Platek et
al. 2003) have been reported to be more
susceptible to yawn contagion than individuals
who score low in those abilities. Consistent
with other empathic behaviors, Norscia and
Palagi (2011) found contagious yawning was more
likely to be elicited in response to the yawning
of more familiar individuals rather than less
familiar individuals. Researchers have shown
particular interest in how the emergence of
contagious yawning across development and
phylogeny can inform us of the underlying
mechanisms of this behavior. Contagious yawning
emerges in children around the same time as more
complex forms of empathy (i.e., 5-6 years of
age, Anderson and Meno 2003; Millen and Anderson
2011) and is disrupted in children with autism
(Giganti and Esposito Ziello 2009; Senju et al.
2007), unless instructed to fixate on the eyes
of the yawner (Senju et al. 2009). Studies with
nonhuman primates further support a possible
association between contagious yawning and
empathy. Chimpanzees (Pan troglodytes) have been
shown to yawn in response to viewing an ingroup
member yawn as opposed to an outgroup member
(Campbell and de Waal 2011), bonobos (Pan
paniscus) exhibit yawn contagion particularly in
response to socially close individuals (Demuru
and Palagi 2012), and gelada baboons
(Theropithecus gelada) exhibit yawn contagion at
higher levels when individuals are socially
close (Palagi et al. 2009). However, yawning in
response to a conspecific's yawn in stumptail
macaques (Macaca arctoides) may be a function of
stress as opposed to empathy, as researchers
observed elevated levels of self-directed
scratching during yawning but not control
stimuli (Paukner and Anderson 2006).
Although it was originally proposed that
complex cognitive abilities (i.e., theory of
mind and self-awareness) are required for the
emergence of contagious yawning (Platek et al.
2003), evidence of contagious yawning in animals
that do not demonstrate these advanced abilities
(e.g., domestic dogs, Canis familiaris)
contradicts this stance. While there are no data
to suggest that dogs possess 'theory of mind,'
dogs are outstanding in their ability to
interpret human social cues. For example, when
given the choice of two containers, one of which
is hiding a piece of food, dogs choose the
correct container at above chance levels after
viewing a human point or gaze at the container
(see review by Reid 2009), outperforming
chimpanzees at this 'objectchoice' task (Hare et
al. 2002). It is possible that through
domestication and coevolution with humans (Hare
et al. 2010), socialization with humans during a
critical period (Udell et al. 2010), or, more
likely, this gene-environment interaction
(Miklo´si and Topa´l 2011), dogs have
developed ways of communicating and coordinating
their behaviors with humans. One means by which
they synchronize their interactions may be
through contagious yawning. However, studies of
yawn contagion in dogs up to this point have
reported variable findings. Although some have
found high levels of contagious yawning in dogs
at the individual level (72 % in Joly-Mascheroni
et al. 2008; 69 % in Madsen and Persson 2012),
others have reported much lower proportions (6 %
in Harr et al. 2009; 26 % in O'Hara and Reeve
2011; 28 % in Silva et al. 2012), possibly
stemming from the considerably different
methodologies employed by each study.
Joly-Mascheroni et al. (2008) suggested two
possible underlying mechanisms for contagious
yawning in dogs. First, dogs may yawn as a means
of coordinating their behavior with humans'
behavior, which may involve an empathic
component. Several studies now have tested the
social modulation of contagious yawning in dogs
(i.e., whether it is biased toward more familiar
individuals, as with other empathic behaviors).
Although Silva et al. (2012) found that dogs
yawned more in response to just the sound of
their owner's yawn than an unfamiliar
individual's, studies with live models have
shown no such relationship (Madsen and Persson
2012; O'Hara and Reeve 2011). In fact, O'Hara
and Reeve found that contactdeprived shelter
dogs yawned more than human-bonded owned dogs,
particularly in response to an unfamiliar human,
supporting Yoon and Tennie's (2010) nonconscious
mimicry hypothesis that contagious yawning may
serve as a means of affiliating with an unknown
human and does not require a 'theory of mind.'
Given the discrepancies in the findings of
previous studies that have tested the social
modulation of contagious yawning, it may be
necessary to use measures that do not rely on
the relationship between the dog and human to
assess the social-cognitive mechanisms that
underlie this behavior.
Alternatively, Joly-Mascheroni et al. (2008)
suggested that contagious yawning in dogs could
be mediated by stress. Mildly stressful
situations may elicit 'tension' yawns in animals
(Maestripieri et al. 1992), particularly dogs
(Beerda et al. 1998). Although tension yawns
resemble 'sleepy' yawns, they often occur with
other stress behaviors such as panting,
trembling, body shaking, lip licking, or
whimpering (Madsen and Persson 2012). Thus,
situations that increase arousal (e.g., the
sound of an owner's yawn in their absence as in
Silva et al. 2012; see Madsen and Persson 2012
for critique) may elicit tension yawns that are
mistaken for contagious yawns. When a live model
is used, the human yawn itself may be
responsible for this increased arousal, which
triggers tension yawns. Joly-Mascheroni and
colleagues proposed that human yawning may be
perceived as a threat behavior to dogs, thereby
causing them to yawn as a means of alleviating
tension, much like what was observed by Paukner
and Anderson (2006) in stumptail macaques.
Although yawning is frequently observed during
agonistic interactions between numerous primate
species (see Baenninger 1997), there is no
scientific evidence that we are aware of to
suggest that human yawning is perceived as a
threat to dogs. Rather, arousal may play an
important role in modulating the meaning of a
yawn, and when displayed in 'situations of
uncertainty' (Madsen and Persson 2012), human
yawning may communicate stress as opposed to
sleepiness to dogs, which leads to an increase
in arousal and subsequent yawning by the dog.
This may also explain O'Hara and Reeve's finding
that contagious yawning was most common in
shelter dogs with unfamiliar humans, since
shelter dogs were more physiological aroused
(measured via heart rate) than owned dogs
throughout testing.
Many of the inconsistencies in the number of
dogs that exhibit yawn contagion presented in
previous studies may be due to the fact that
different methodologies produce varying levels
of arousal in dogs (Madsen and Persson 2012).
Most studies attempt to control for tension
yawns through methodological procedures (e.g.,
testing in a familiar environment) and excluding
yawns that occur in the presence of other stress
behaviors. Although some have presented
convincing evidence for contagious yawning in
dogs by taking these measures (e.g., Madsen and
Persson 2012), it is still unclear how these
methodologies influence arousal (O'Hara and
Reeve 2011; Silva et al. 2012). Because there is
great individual variation in dogs' behavioral
coping strategies in response to stressors
(Beerda et al. 1998), physiological measures are
needed to determine the role arousal is playing
in contagious yawning in dogs. Many have
recognized the need for physiological measures
when assessing contagious yawning in dogs
(Joly-Mascheroni et al. 2008; Madsen and Persson
2012), but only one study up to this point has
used a physiological measure to quantify stress
responses in their subjects (i.e., heart rate:
O'Hara and Reeve 2011), and the impact that
testing had on individual dogs' physiological
responses and subsequent yawning was
unclear.
The purpose of the present study was to test
the two explanations put forth by
Joly-Mascheroni et al. (2008) in order to
determine why some dogs exhibit yawn contagion
in response to human yawns while others do not.
We examined the role of arousal in contagious
yawning using a physiological measure that has
not yet been used in yawn contagion studies.
Specifically, we assessed whether dogs that
exhibited yawn contagion had elevated levels of
arousal via salivary cortisol measures. We also
included a measure to assess whether
social-cognitive processes play a role in
contagious yawning in dogs as in some primates
(Platek et al. 2003; Campbell and de Waal 2011).
We propose that if yawning serves as a means of
coordinating interactions between dogs and
humans, perhaps dogs that exhibit yawn contagion
are also more perceptive to human social cues in
general. Based on previous studies that have
shown that social-cognitive performance (i.e.,
faux pas theory of mind and self-face
recognition tasks) correlates with contagious
yawning in humans (Platek et al. 2003), we
expected that dogs that exhibited yawn contagion
would also perform better on a social-cognitive
task. We chose to test these two mechanisms
using shelter dogs in order to elaborate on
O'Hara and Reeve (2011) study in which they
unexpectedly found that shelter dogs exhibited
yawn contagion at much higher levels than owned
dogs.
Discussion
This study is the first to present evidence
to suggest that human yawning, when presented in
a stressful context, influences physiological
arousal and yawning in some dogs. Although we
did not observe contagious yawning among shelter
dogs overall, we identified 12 dogs that yawned
only in the yawning condition (henceforth
referred to as dogs that appeared to exhibit
yawn contagion). This subgroup of dogs was
distinct from the other dogs in this study in
their levels of physiological arousal following
exposure to human yawns. Specifically, their
cortisol levels remained elevated following
exposure to human yawning, whereas other dogs
had reduced levels relative to their baseline
levels. We must caution that we did not observe
notable differences between the number of dogs
that yawned only in the yawning condition versus
the control condition and that it is possible
that some dogs may have fallen into this
category by chance. However, if a significant
number of these dogs were in this subgroup by
chance, it is unlikely that we would have
observed these different cortisol patterns.
Further, this effect did not appear to be the
result of some dogs being more inherently
nervous, causing them to yawn more, because
there was no relationship between baseline
cortisol levels and yawning rates.
In addition to different cortisol patterns,
83 % of dogs that yawned contagiously had
received the yawning stimuli rather than control
stimuli first. Previous studies would support
that levels of arousal were probably elevated in
the first trial regardless of condition as a
result of the novelty of the situation (Tuber et
al. 1996; Hennessy et al. 1998). One possibility
is that when yawning is presented in the first
trial, when arousal is high, these dogs are more
likely to yawn and their cortisol levels remain
elevated, but when yawning stimuli are presented
later on, after dogs have acclimated and arousal
is lower, they are less likely to yawn. It is
also possible that dogs that received control
stimuli first habituated to mouth movements,
which decreased their responsiveness in the
second trial when yawning stimuli were presented
and therefore showed lower levels of yawning.
However, since our findings suggest that
elevated arousal may facilitate yawning, and
yawning has been directly associated with
glucocorticoids in previous studies
(An�´as-Caldero´n et al. 2004), we
tend to agree with the first
interpretation.
These findings suggest that context and
arousal could both play a role in modulating the
meaning of a yawn. Previous studies have shown
that shelter dogs have elevated cortisol levels
relative to pet dogs in their homes, especially
during the first few days in the shelter
(Hennessy et al. 1997). We suggest that when a
yawn is presented in situations of uncertainty,
when arousal is high (e.g., in a shelter
environment), it may be nonconsciously perceived
as a signal for stress, but when presented in a
comfortable situation, when arousal is low
(e.g., in a home environment), it may be
nonconsciously perceived as a signal for
sleepiness. The perception of the yawn then
influences the arousal state of the perceiver
(i.e., the dog) and the type of yawn that is
exhibited (tension yawn or true contagious
yawn). However, the direction of the
relationship between arousal and a yawning
response is unclear. It has been suggested that
in true, 'sleepy' contagious yawning, the
yawning response serves as a low-level form of
mimicry (Yoon and Tennie 2010) that is then
internalized and produces a reduction in arousal
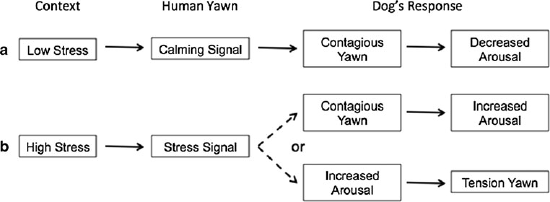
(Madsen and Persson 2012; Fig. 4a). This
suggestion has been supported by anecdotal
evidence of diminished arousal in dogs following
yawning, but not control stimuli presented in a
naturalistic, comfortable setting (Madsen and
Persson 2012). It is possible that the yawns we
observed in this study follow this process
(i.e., dogs mimic the yawn of the actor, and
given the heightened arousal of the dog and the
uncertain context, dogs show an increase in
arousal). Like true contagious yawning, this
would also serve to synchronize their behavior
with humans' (Joly-Mascheroni et al. 2008).
However, since nearly an equal number of dogs
yawned only in the control condition as in the
yawning condition, our data do not suggest that
dogs were mimicking the human's behavior. On the
other hand, it may be more plausible that a
human yawn could trigger a physiological
response (i.e., elevated arousal), which then
elicits tension yawns. Rather than functioning
to coordinate behaviors, these yawns would serve
to displace internal anxiety arising from a
tense social situation (Maestripieri et al.
1992; Fig. 4b). Whether this is in fact the case
could be determined in future studies by
blocking physiological arousal and observing
whether dogs in high stress contexts continue to
yawn.
Even though the evoked yawns that occur in
high arousal situations, like those observed in
this study, may differ from true contagious
yawns, they could potentially still involve a
social-cognitive mechanism. Human yawning
influenced dogs' levels of arousal, which
suggests that the perception of a human yawn
contains a communicative component, perhaps
conveying their emotional state. This may also
be related to dogs' ability to empathize with
humans. However, recent studies have reported
variable findings as to whether contagious
yawning in dogs is biased toward familiar
individuals (Madsen and Persson 2012; O'Hara and
Reeve 2011; Silva et al. 2012). Unlike primates,
which have demonstrated social modulation of
contagious yawning (e.g., bonobos: Demuru and
Palagi 2012; chimpanzees: Campbell and de Waal
2011, but see Massen et al. 2012; humans:
Norscia and Palagi 2011; gelada baboons: Palagi
et al. 2009), dogs may not, possibly because of
their frequent positive interactions with
unfamiliar humans (Campbell and de Waal 2011),
or perhaps the emotional bond between human and
dog is only unidirectional (O'Hara and Reeve
2011). Before concluding that this lack of
social modulation suggests that contagious
yawning operates on a low-level form of
nonconscious mimicry or affective empathy
(Madsen and Persson 2012), we should generate
alternative means of assessing the underlying
social-cognitive mechanisms of contagious
yawning.
Rather than exploring a familiarity bias to
determine the role of social cognition in yawn
contagion, we explored whether dogs that
exhibited yawn contagion were exceptional in
their social-cognitive abilities, similar to the
premise of Platek et al. (2003) study with
humans. The objectchoice task involves a basic
level of perspective-taking (Emery and Clayton
2009), which is an important skill for
coordinating group behavior. The object-choice
task was chosen to assess the social-cognitive
skills of the dogs in this study because it
encompasses the basic components of more complex
abilities that have been suggested to be
involved in yawn contagion (Platek et al. 2003).
However, we found that dogs that exhibited yawn
contagion in this study performed similarly on
the object-choice task to dogs that did not yawn
uniquely at the yawning stimuli. There are
several ways to interpret our finding that
social-cognitive performance was not different
among dogs that did and did not yawn
contagiously. First, this may suggest that yawn
contagion may not require the high level of
cognitive complexity (i.e., theory of mind) that
had been proposed by Platek et al. (2003). This
would support Madsen and Persson's (2012)
suggestion that lower-level affective empathy
underlies contagious yawning in dogs rather than
more complex cognitive empathy. However, we also
must acknowledge that the seemingly 'contagious'
yawns we observed in this study may operate on
some different mechanism than the contagious
yawns observed in other studies (e.g., Madsen
and Persson 2012), given our subjects'
heightened arousal compared to owned dogs
(Hennessy et al. 1997). This could also explain
why dogs that exhibited yawn contagion performed
the same as other dogs on the object-choice
task. Further, dogs that yawned in response to
human yawns had elevated levels of physiological
arousal, and heightened arousal was associated
with poorer performance on the object-choice
task. Thus, our measure of social-cognitive
abilities may have been confounded by high
levels of arousal. It may be necessary to test
owned dogs in their homes under more comfortable
conditions (as in Madsen and Persson 2012) in
order to determine whether dogs that exhibit
yawn contagion in comfortable settings are in
fact more perceptive to human social cues. If we
still fail to observe an association between
performance on the object-choice task and yawn
contagion when tested in a comfortable setting,
this would not necessarily imply that social
cognition does not underlie contagious yawning
in dogs. Rather, it may indicate that this type
of social-cognitive task is assessing a process
unrelated to contagious yawning. Still,
researchers should strive to develop other ways
of testing the relationship between yawn
contagion and social cognition (including
empathy) aside from familiarity biases.
Though some studies have provided convincing
evidence that dogs engage in yawn contagion with
humans (e.g., Joly-Mascheroni et al. 2008;
Madsen and Persson 2012), other studies have
found only a small percentage of dogs exhibits
this behavior (Harr et al. 2009; O'Hara and
Reeve 2011). The discrepancies between these
studies may be the result of the fact that yawn
contagion in dogs, though present, is influenced
by contextual factors more so than in other
species (i.e., primates, Demuru and Palagi
2012). Our findings indicate that in a stressful
context, dogs that yawn in response to human
yawns remained aroused following exposure to
stimuli. Though these yawns appear contagious,
they may be based on different underlying
mechanisms than what would be observed in a
low-stress situation. It is possible that the
methods utilized in this and other studies
(e.g., requiring eye contact, long periods of
silence with little if any petting, testing in
an unfamiliar setting) may have influenced
arousal levels and therefore could contribute to
the variable findings between studies. This
emphasizes the importance of assessing arousal
in such studies. Potential stressors (e.g.,
unfamiliar humans, novel environment), including
stimuli that may trigger physiological arousal
that may not typically be considered a
'stressor' (e.g., food, exercise, eye contact),
should be considered in their contribution to
dogs' state of arousal.
Further, as there is no standardized
behavioral measure of stress in dogs given the
individual variation in behavioral
manifestations of stress (Rooney et al. 2007),
physiological measures can provide valuable
information regarding the role of physiological
arousal in contagious yawning. In fact,
physiological measures may be necessary in
situations in which it is not clear whether the
contagious yawning being observed is the result
of arousal and tension yawns.
Conclusions
Our findings are consistent with several
previous studies that reported low levels of
contagious yawning in dogs. However, these
results may be explained by the arousal states
of our subjects. Dogs that exhibited yawn
contagion remained physiologically aroused
following observing human yawns, and most of
them had received the yawning stimuli first.
These findings may suggest that arousal levels
and context have a substantial effect on the
perception of a human yawn by dogs. When
presented in conjunction with other
environmental stressors, human yawning may be
perceived as a signal for stress, leading to
increased arousal levels, which then elicit
tension yawns. Although these yawns resemble
true contagious yawning, they probably operate
on some other mechanism. Investigating the
socialcognitive mechanisms that underlie this
phenomenon can reveal whether this type of
yawning is functionally distinct from contagious
yawning observed in other studies when arousal
is low. Although we did not find that the type
of contagious yawning we observed in this study
was linked to dogs' social-cognitive abilities,
it is important to explore alternative means of
assessing social cognition and empathy when
attempting to understand the processes that give
rise to contagious yawning in dogs.