Abstract : Our ability to explain and
predict other people's behaviour by attributing
to them independent mental states, such as
beliefs and desires, is known as having a
'theory of mind'. Interest in this very human
ability has engendered a growing body of
evidence concerning its evolution and
development and the biological basis of the
mechanisms underpinning it. Functional imaging
has played a key role in seeking to isolate
brain regions specific to this ability. Three
areas are consistently activated in association
with theory of mind. These are the anterior
paracingulate cortex, the superior temporal
sulci and the temporal poles bilaterally.
This review discusses the functional
significance of each of these areas within a
social cognitive network.
The role of the STS
In addition to the anterior paracingulate
cortex, two regions, the STS and the temporal
poles bilaterally, consistently activate in
studies of theory-of-mind ability. This activity
is probably not related to the decoupling that
is necessary for mentalizing. The functions of
these regions might relate to abilities that aid
mentalizing. It is from these pre-existing
abilities that mentalizing has evolved. The
precise role of the STS is still unclear.
Gallagher et al. found predominantly right STS
to be associated with understanding the meaning
of stories and cartoons involving people, with
or without the requirement to mentalize. Other
functional neuroimaging studies of
theory-of-mind tasks have associated STS
activity with the involvement of people when
understanding causality and intentionality, the
attribution of intentions to the movements of
geometric shapes and taking the
self-perspective. It is likely that all of these
tasks share one common function that is able to
account for this consistent activation.
Biological motion
Functional imaging studies that examine the
perception of biological motion activate the
same region of the STS in association with hand
actions, body movements, mouth movements and lip
reading, implied biological motion and eye
movements and gaze direction. In addition,
Narumoto et al. found that the right STS was
part of a network of face-responsive brain
regions. Selective attention to facial emotion
specifically enhanced the activity of the right
STS compared with attention to the face per se.
They attribute this activation to facial emotion
recognition within a distributed face-processing
system. Consideration of these findings has led
researchers to speculate on the role of this
region within a social cognitive network.
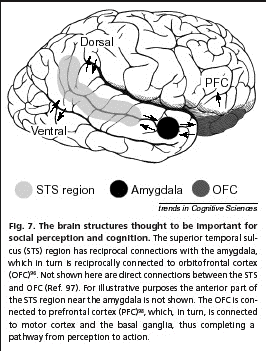
Allison et al. suggest that the STS is sensitive
to stimuli that signal the actions and
intentions of another individual. The results of
neuroimaging, electrophysiological and
single-cell recording studies converge to
suggest that initial analysis of social cues
occurs in the STS region, which is anatomically
well sited to integrate information derived from
both the ventral and dorsal visual pathways. In
accordance with this notion, Frith and Frith
suggest that the STS is involved in the
detection of the behaviour of agents and
analysis of the goals and outcomes of this
behaviour. The results of the Gallagher et
al.study, in particular, indicate that the right
STS is involved in explaining the behaviour of
others as a result of recognizing a physical
cause or their mental state. This idea gains
support from a recent fMRI study of social
judgement. Winston et al. asked volunteers to
view faces and make judgements on either
trustworthiness or age. They found right STS
activity when volunteers made explicit
judgements about trustworthiness. The authors
attribute this activity to intention detection
from visual cues, a critical component in
determining whether or not to trust an
individual.
The role of the temporal poles
The temporal poles are generally associated
with object and face recognition in primates.
However, functional imaging studies of humans
have activated the temporal poles in association
with the broader context of episodic memory
retrieval in visual and auditory domains. Thus
the temporal poles are active during the
recollection of familiar faces and scenes, the
recognition of familiar voices, emotional memory
retrieval and autobiographical memory retrieval.
Taken together, these results suggest that the
temporal poles are a store for personal semantic
and episodic memories.
There are several reasons why episodic
memory might be useful for mentalizing. If we
are engaged in deception we need to remember
what we said to someone the last time we met
them. We might remember past episodes in which
the behaviour we are observing now was
associated with a particular mental state.We
might draw on our past experience to imagine
ourselves in the situation of another person and
thereby 'simulate' their experience. We have
already mentioned the importance of semantic
memory for generating the scripts that enable us
to anticipate the likely goals of particular
people in particular situations. Patients with
semantic dementia show atrophy in the anterior
temporal lobes, especially on the left. As this
atrophy progresses, these patients lose
knowledge of all but the simplest and most
concrete scripts. This should lead to specific
difficulties with theory-of-mind tasks.