Mutschler I, Reinbold C, Wankerl J, Seifritz
E, Ball T.
Abstract
Empathy is key for healthy social
functioning and individual differences in
empathy have strong implications for manifold
domains of social behavior. Empathy comprises of
emotional and cognitive components and may also
be closely linked to sensorimotor processes,
which go along with the motivation and behavior
to respond compassionately to another person's
feelings. There is growing evidence for local
plastic change in the structure of the healthy
adult human brain in response to environmental
demands or intrinsic factors. Here we have
investigated changes in brain structure
resulting from or predisposing to empathy.
Structural MRI data of 101 healthy adult females
was analyzed. Empathy in fictitious as well as
real-life situations was assessed using a
validated self-evaluation measure. Furthermore,
empathy-related structural effects were also put
into the context of a functional map of the
anterior insular cortex (AIC) determined by
activation likelihood estimate (ALE)
meta-analysis of previous functional imaging
studies. We found that gray matter (GM) density
in the left dorsal AIC correlates with empathy
and that this area overlaps with the domain
general region (DGR) of the anterior insula that
is situated in-between functional systems
involved in emotion-cognition, pain, and motor
tasks as determined by our meta-analysis. Thus,
we propose that this insular region where we
find structural differences depending on
individual empathy may play a crucial role in
modulating the efficiency of neural integration
underlying emotional, cognitive, and
sensorimotor information which is essential for
global empathy.
Introduction
Empathy has strong implications for manifold
domains of social behavior and it may constitute
an integral part of emotional intelligence
(Goleman, 1998). In the psychological
literature, empathy has been defined as the
ability to feel what another person is feeling
(emotional component), and knowing what a person
is feeling (cognitive component), i.e., to
achieve a cognitive understanding of other
feelings (Eisenberg and Miller, 1987; Decety and
Jackson, 2004). Empathy may also include
sensorimotor processes, which go along with the
motivation and behavior to respond
compassionately to another person's feelings
(Preston, 2007; Zaki and Ochsner, 2012). It has
been shown that empathetic ability, however, is
not always going along with prosocial behavior
(Eisenberg and Miller, 1987).
The cognitive component of empathy may be
closely linked to "theory of mind," that is the
meta-cognitive ability to represent mental
states such as beliefs, intentions, and desires
of other people (Premack and Woodruf, 1978).
According to Dymond, an empathic person can
imaginatively take the role of another and can
understand and predict that person's thoughts,
feelings, and actions (Dymond, 1949). Together,
these definitions suggest that the human ability
of empathy is more than a pure emotional process
but also includes cognitive aspects such as
perspective taking and may also involve the
sensorimotor system for responding appropriately
and compassionately to another person's
feelings. There is growing appreciation that
there are stable differences between individuals
in the level of empathy which has a high impact
on social behavior (Zaki and Ochsner, 2012) and
that these differences can be reliably measured
(Miller and Eisenberg, 1988; Marangoni et al.,
1995; Singer et al., 2004).
Recent studies have shown that there is
capacity for local plastic change in the
structure of the healthy adult human brain in
response to environmental demands or intrinsic
factors (Johansen-Berg, 2012). There is
increasing interest in investigating the
neuroanatomical basis underlying individual
differences in human behavior and cognition
(Kanai and Rees, 2011), including empathy
(Banissy et al., 2012). However, the structural
basis of empathy in particular in the insular
cortex has remained unclear. Accumulating
evidence indicates a crucial role of the insular
cortex in empathy: in particular the anterior
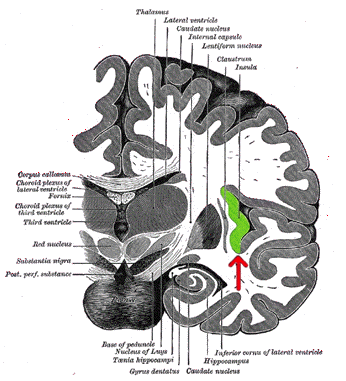
insular cortex (AIC)a brain region which is
situated in the depth of the Sylvian fissure and
anatomically highly interconnected to many other
cortical regions (Nieuwenhuys, 2012) is part of
the functional neural network that plays an
essential role in mediating social-emotional
processing (Mutschler et al., 2009, 2012)
including empathy (Singer et al., 2004; Seeley
et al., 2012).
Discussion
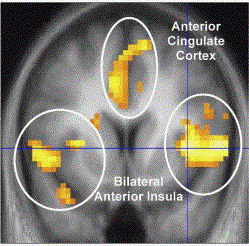
Our results indicate regionally specific
structural differences in the left dorsal AIC
related to individual empathy scores in healthy
individuals. GM density correlated in a confined
region of the left dorsal AIC with empathy in
healthy females. The insular region where we
find structural differences overlaps with the
peaks from a previous functional study
correlating empathy-related responses fMRI with
individual empathy scores (Singer et al., 2004),
Figure 2B. This precise spatial correspondence
strongly supports our VBM findings. Studies show
that individuals with specialized abilities have
markedly developed brain structures in specific
regions related to their expertise (Kanai and
Rees, 2011). For instance, an investigation by
Maguire et al. on London taxi drivers found that
GM measures derived from T1-weighted structural
MRI are sensitive to experience. The authors
showed that taxi drivers, with their knowledge
of London's complex street plan, had enlarged
posterior hippocampi in comparison to control
subjects (Maguire et al., 2000). More recently,
a longitudinal study demonstrates that learning
to juggle increases GM density in
occipito-parietal cortical areas in the adult
brain involved in reaching and grasping (Scholz
et al., 2009).
The results reveal that our morphometrically
identified area in the dorsal AIC related to
individual differences in empathy overlaps the
DGR. A first indication toward a DGR was
provided by Dosenbach et al. who conjointly
analyzed mixed design fMRI experiments using 10
different tasks (Dosenbach et al., 2006). They
found, among others, the bilateral anterior
insula/frontal operculum region to show reliable
start-cue and sustained activations across all
or nearly all tasks. This generalized type of
activation was found in the dorsal part of the
AIC (black dot in Figure 2). The idea of a DGR
was extended by Craig in his review on awareness
and the AIC (Craig, 2009).
The author observed that, in imaging
studies, the AIC is reported to be activated in
an astonishing number of studies from a broad
range of topics including all types of
subjective feelings, attention, cognitive
choices, intentions, music, time perception,
awareness of sensations, and movements, of
visual and auditory percepts, of the
trustworthiness of other individuals and
concluded that "No other region of the brain is
activated in all of these tasks." In a
subsequent meta-analysis of a wide range of
functional imaging studies, the same region of
the AIC that showed task-set-related responses
in the study by Dosenbach et al. was also
reliably activated by a wide range of auditory
and language tasks as well as during social norm
violation (Mutschler et al., 2009). Importantly,
this study was restricted to studies which
reported clear insular responses, i.e.,
ambiguous effects such as "insula/frontal
operculum" were excluded, giving strong
additional support for the existence of a
domain-general region in the AIC. The idea of a
DGR that is activated across nearly all kinds of
tasks as well as Craig's observation that no
other brain region shows such generalized
responses was later also confirmed in a
meta-analysis (Kurth et al., 2010).
However, little is known regarding the exact
location and the functional meaning of the DGR
in the dorsal AIC. There is good evidence that
the DGR is found in the dorsal anterior insula
as indicated in Figure 2A. Importantly, compared
to a previous interpretation (Brown et al.,
2011) our ALE analysis shows that
movement-related responses do not (or only to a
very small degree) overlap with the
domain-general area. The position of the main
region with reproducible movement-related
responses is located just posterior to the
domain-general region (blue area in Figure 2).
This movement-related area closely co-localized
with insular peaks related to the sense of
agency of hand movements (Mutschler et al.,
2009). What is the functional meaning of the
DGRin particular for empathy? Several
overarching functions have been proposed such as
task-set processing (Dosenbach et al., 2006) and
a role in awareness (Craig, 2009). Importantly,
our findingsthat the morphometrically identified
area in the dorsal AIC related to individual
differences in empathy overlaps the DGRsupport
the notion that its underlying neuronal
substrate may be involved in integrating
socio-emotional information during empathy.
We find that emotion-related responses in
healthy subjects were preferentially located in
the dorsal AIC. This region was distinct from
the insular region associated with peripheral
physiological changes resulting from emotional
experiences and found that this activity was
represented in the ventral AIC that was also the
most likely site of insular co-activation with
the amygdala (Mutschler et al., 2009). In the
current study, insula-coordinates associated
with emotion-related peripheral physiological
changes were excluded. Therefore, our findings
suggest that distinct functional insula regions
may be involved in different aspects of
emotional processing, whereby peripheral
physiological correlates of emotional processing
are mapped to the ventral anterior regions,
while emotion-cognitive processes are mapped to
a more dorsal-anterior region. More
specifically, we assume that the dorsal anterior
insula might play a pivotal role in integrating
sensory stimuli with its salience, possibly via
connections to the cingulate cortex. This
interpretation is supported by the fact that in
our study-sample subjects evaluated the
emotional content of the presented stimuli and
in direct agreement with recent findings showing
that the dorsal anterior insula is more
consistently involved in human cognition than
ventral anterior and posterior networks (Chang
et al., 2013), and the observed functional
connectivity between the dorsal anterior insula
and the dorsal anterior cingulate cortex, which
that plays a crucial role in cognitive
decision-making (Deen et al., 2011).
Pain-related maximal ALE were found in the
posterior insula and in the dorsal AIC. Both,
emotion and pain related ALE findings are
discussed in more detail elsewhere (Mutschler et
al., 2012) because the aim of this meta-analysis
was to relate them to studies on empathy.
Recent neuroimaging studies show that the
anterior insula and the anterior cingulate
cortex are engaged during both, the experience
and observation of pain (Singer et al., 2004;
Lamm et al., 2011). It has been suggested that
neural responses involved in both conditions
might reflect a neuronal substrate which is
related to the affective but not sensory aspect
of pain (Singer et al., 2004). Together, our
present ALE-findings support this notion and
suggest that the posterior insula is involved in
processing sensory aspect of pain, whereas the
dorsal anterior insula is involved in emotion.
In summary, we argue that the dorsal AIC plays a
pivotal role in empathy (similarly as during
emotion processing and pain) by integrating
sensory stimuli with its salience, possibly via
connections to the cingulate cortex. This
assumption is also supported by the fact that
ALE-findings related to emotion and empathy for
pain and also the DGRwhich has been associated
with cognitionoverlap in the dorsal anterior
insula, suggesting that these functions share a
common neural substrate (Dosenbach et al.,
2006). As mentioned above we assume that the
overall role of the morphometrically identified
area in the dorsal AIC related to individual
differences in empathy which overlaps the DGR
might be involved in integrating information
which is relevant for socio-emotional and
cognitive processing. Thus, we assume that
empathy is not (only) related to a specific
"socio-emotional" interaction area, but to a
superordinate "domain-general" area, in line
with concepts of empathy that include not only
social and emotional, but also cognitive aspects
(Eisenberg and Miller, 1987; Decety and Jackson,
2004). Whether our findings in the dorsal AIC
have also a relation to the "von Economo
neurons" [VENs, (Von Economo, 1926)]
remains to be determined. VENs have been
hypothesized to play are role in
social-emotional processing including empathy
(Evrard et al., 2012; Seeley et al., 2012).
In the following paragraph potential
limitations of this study are discussed and
suggestions are made for future research.
Similarly to previous imaging studies (e.g.,
Singer et al., 2004) we have investigated
correlates of overall empathy in our study by
using a widely applied and validated
self-evaluation measure (Leibetseder et al.,
2001, 2007). There is a potential concern about
the influence of the social desirability
biaswhich refers to the tendency of subjects to
answer self report items in a self-favoring
manneron the validity of questionnaire-based
research (Edwards, 1957). It is discussed
whether social desirability scales may be used
to detect, minimize, and correct for social
desirability bias in order to improve the
validity of questionnaire-based research (e.g.,
Uziel, 2010). In a future study it would be
interesting to measure empathy
experimentallye.g., by investigating the impact
of compassion-based meditation on empathy
(Mascaro et al., 2013)and to relate functional
activity to GM properties.
In this study, brain structure changes
resulting from or predisposing to empathy have
been investigated in a large sample of females
because of increasing evidence for sex
differences in empathy. Females score on average
higher than males on self-report measures of
empathy (Hoffman, 1977; Baron-Cohen and
Wheelwright, 2004). Singer et al. observed in an
fMRI study on empathy for pain that in males but
not females empathetic reactions were absent for
persons who were perceived as behaving unfairly
(Singer et al., 2006). Recently, Van Honk et al.
(2011) showed that the administration of the
androgen hormone testosteronewhich represents
the largest hormonal difference between the
sexesimpaired cognitive empathy in healthy
females. As mentioned, the insular region where
we find structural differences exactly overlaps
with the peaks from a previous functional study
correlating empathy-related responses fMRI with
individual empathy scores (Singer et al., 2004).
Notably, in this study also only females were
investigated. A recent study found in a sample
with mixed gender individual differences in
trait empathy dimensions correlating with
morphological differences in several brain areas
including the anterior cingulated cortex and the
right dorsolateral prefrontal cortex (Banissy et
al., 2012). More specifically, based on an
analysis with ROIs around peaks from previous
functional studies on empathy, Banissy et al.
report structural changes in the ventral-most
part of the insular cortex, several centimeters
apart from the area characterized in the present
study.
Interpretational difficulties however arise
because (1) the peak at MNI coordinates -39, 9,
-21 as reported by Banissy et al. (that was used
to define the ventral insular ROI) is
pain-related and not empathy-related as was
assumed by the authors (see Appendix to Singer
et al., 2004) and (2) a second, empathic
concern-related peak reported to be in the
insular region by Banissy et al. at MNI
coordinates -48, 6, 18 is according to the
probabilistic assignment obtained from the SPM
Anatomy Toolbox (Eickhoff et al., 2005) located
in Brodmann Area 44. In this study, structural
changes specific to males or females were
however not addressed and only changes that were
consistent across the whole (mixed) sample were
reported which may possibly explain the
different results of their and our study.
Alternatively, the different results between
both studies may also be explained by the fact
that in our study the focus was on global
empathy whereas in the study by Banissy et al.
on the relationship between components of
empathy (empathic concern, personal distress,
perspective taking, and fantasizing) and brain
structure using a different measure. Because
empathy crucially requires high-level
integration of emotional, cognitive, and social
components as well as of behavioral control the
goal of our study was not to investigate
different components of empathy but to explore
the neural substrate that may underlay its
neural integration. Our interpretationthat the
dorsal AIC where we find structural differences
depending on individual empathy might play a
crucial role in modulating the efficiency of
neural integration underlying emotional,
cognitive, and sensorimotor information which is
essential for global empathy is in agreement
with previous studies reporting that reduced GM
volume in the AIC was associated with a lack of
empathy in neuropsychiatric disorders such as in
conduct disorder (Sterzer et al., 2007) and in
psychopathy (de Oliveira-Souza et al., 2008).
Further, as mentioned above, a recent study in
combat veterans with traumatic brain injury
shows that lesions in several brain regions,
particularly in the insula, was associated with
deficits in empathy (Driscoll et al., 2012). In
future research based on larger samples it would
be interesting to investigate the role of
empathy subscales such as cognitive and
emotional components (Leibetseder et al., 2007),
and their relation to functional and structural
data. The emotional component of empathy has
been for instance closely linked to activation
in the inferior frontal gyrus (Shamay-Tsoory et
al., 2009; Banissy et al., 2012).
Finally, longitudinal studies are needed to
clarify whether the empathy-related structural
effects in the dorsal AIC that we find are due
to a pre-existing brain characteristic or to
empathy-experience, or whether it indicates
both.
In addition, all neuroimaging studies
entering our meta-analysis reported that only
healthy individuals free of any neurological and
psychiatric disorders were investigated.
However, for future meta-analyses it would be
important that neuroimaging studies specify more
clearly the procedure regarding how they
assessed and excluded individuals with
neurological and psychiatric disorders
[e.g., whether a structured clinical
interview for Diagnostic and Statistical Manual
IV (DSM-IV) axis I and axis II personality
disorders was used]. Furthermore, it is also
important to note that our meta-analysis on
empathy included females and males. An
interesting topic for future meta-analyses would
be to examine whether there are functional
differences between male and female samples with
respect to empathy. Finally, future
meta-analyses should also investigate whether
the dorsal AIC is rather involved in empathy for
pain (Singer et al., 2004) and social rejection
(Eisenberger et al., 2003) than in empathy for
positive emotions and if yes why this could be
the case. Only few brain imaging studies to date
have examined empathy for positive emotions
(Jabbi et al., 2007; Mobbs et al., 2009; Morelli
et al., 2012), therefore ALE meta-analysis could
not be performed. If the dorsal AIC is related
to empathy and is essential for high-level
integration, it should be activated during
empathy for all types of emotions. However, the
few studies on empathy for positive emotions
suggest that the dorsal AIC might not be
involved (Mobbs et al., 2009; Morelli et al.,
2012), but a meta-analytic analysis based on a
large sample of studies would be required to
resolve this issue. In future it would be also
important to meta-analytically examine studies
on empathy for negative emotions which show
insula activation, such as empathy for disgust
(Wicker et al., 2003), embarrassment (Krach et
al., 2011), social rejection (Masten et al.,
2011), and anxiety (Morelli et al., 2012), as
well as studies on functional components of
empathy.
In summary, the dorsal AIC where we
find structural differences depending on
individual empathy may be key in modulating the
efficiency of neural integration underlying
emotional, cognitive, and sensorimotor
information which is essential for empathy.
Furthermore, our results support a functional
subdivision of the human insula in functionally
distinct regions. These include the ventral
anterior insula which is involved in mapping
peripheral physiological information during
emotional experiences and the dorsal AIC which
plays a crucial role in integrating sensory
stimuli with salience possibly via connections
to the cingulate cortex. The dorsal anterior
insula constitutes an auditory and language area
and the mid anterior insula plays a pivotal role
in sensorimotor processing. Finally, the
posterior insula may be involved in processing
sensory aspects of nociceptive information and
the dorsal AIC may have an integrative role
during emotional-cognitive evaluation of a
noxious stimuli and the associated sensorimotor
response. Together, these findings provide new
important insights into the functional
organization of the human insular cortex in
healthy individuals, and the functional map may
be helpful for understanding dysfunction in
conditions affecting empathy such as borderline
personality disorder with co-morbid
posttraumatic stress disorder, autistic spectrum
disorders, psychopathy/antisocial personality
disorder, conduct disorder, and
schizophrenia.