Department of Pharmacology,
Fukuoka University, Japan
Introduction
Systemic administrations of low doses of
dopamine receptor agonists such as apomorphine,
bromocriptine and quinpirole have been reported
to elicit yawning in rats. The yawning behavior
induced by dopamine receptor agonists is blocked
by both dopamine D2-receptor and muscarinic
receptor antagonists. Previous investigations
have also pointed out the involvement of the
cholinergic system in yawning behavior.
Physostigmine, an anticholinesterase agent, and
pilocarpine, a muscarinic receptor agonist,
induce a yawning behavior which is inhibited by
muscarinic receptor antagonists. On the basis of
such findings, it has been proposed that yawning
seems to involve dopamine D2receptor stimulation
and/or muscarinic receptor activation.
The secretion of prolactin in rats is
regulated by neurotransmitters and hypothalamic
hormones which have inhibitory or stimulatory
effects. Especially, the tuberoinfundibular
dopamine system is proposed to participate
mainly in the regulation of prolactin secretion.
Actually, serum prolactin levels are decreased
by bromocriptine, a dopamine D2-receptor
agonist, and increased by haloperidol, a
dopamine receptor antagonist.
Recently, new types of dopamine receptor
agonists have been introduced.
6-Allyl-2-amino-5,6,7,8-tetrahydro-4Hthiazolo
[4,5-d] azepine (talipexole, B-HT 920)
is characterized as an agonist for brain
dopamine autoreceptors. This drug is found to
induce yawning and to decrease basal prolactin
levels in rats.
(S)-2-amino-4,5,6,7-tetrahydro-6-propylamino-benzothiazole
(SND 919) is most recently synthesized and is
expected to be a compound possessing
talipexole-like dopamine receptor agonistic
activities and fewer alpha-adrenoceptor
stimulating effects.
Therefore, the aim of the present study was
to investigate whether SND 919 induces yawning
and decreases prolactin secretion via
stimulation of dopamine D2-receptors.
Discussion
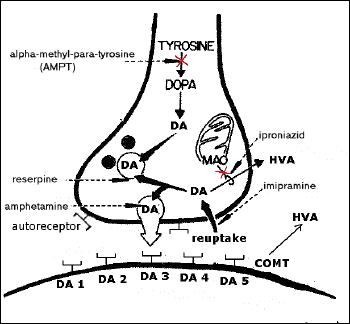
Biochemical and pharmacological evidence
indicates the presence of at least two dopamine
receptor subtypes; dopamine D1-receptors linked
positively to adenylate cyclase and dopamine
D2-receptors not linked or linked negatively to
adenylate cyclase. The present study shows that
SND 919 as well as talipexole possesses a
selective dopamine D2-receptor agonistic
property in the rat brain and the anterior
pituitary gland. It has been proposed that
talipexole does not exert any postsynaptic
dopaminergic effects such as locomotor
hyperactivity and stereotyped behavior in naive
animals with normosensitive brain dopamine
receptors. However, recent observations have
shown that talipexole also exhibits postsynaptic
dopamine receptor agonistic properties under
pretreatment with the dopamine D1-receptor
agonist, SK&F 38393. This combined treatment
with talipexole and SK&F 38393 induced a
strong stereotyped behavior in either naive or
reserpine plus alpha-MT-pretreated rats. This
effect was analogous to the postsynaptic effect
under high doses of apomorphine, a mixed
dopamine D1/D2-receptor agonist. Thus, it has
been proposed that talipexole is a selective
dopamine D2-receptor agonist and that its own
postsynaptic agonistic effects are masked in
animals with normosensitive postsynaptic
dopamine D2-receptors by its particularly potent
action on the dopamine autoreceptors and
consequent critical lack of synaptically
available dopamine.
On the other hand, yawning is reported to be
evoked, without eliciting stereotyped behavior,
with low doses of dopamine receptor agonists
such as apomorphine, piribedil, (+)-3-PPP,
quinpirole or bromocriptine. The yawning induced
by dopamine receptor agonists is also blocked by
dopamine D2-receptor antagonists and muscarinic
receptor antagonists. Actually, in the present
experiment, low doses of SND 919, talipexole or
(+)-3-PPP induced a marked yawning in naive rats
without producing any stereotyped behaviors.
Moreover, the administration of SND 919 in low
doses (50, 100 .ig/kg, s. c.), which could
induce yawning, failed to elicit stereotypy in
the rats treated with SK&F 38393 (10 mg/kg,
s. c.), but high doses (500, 1000 pg/kg, s.c.)
of the drug did evoke stereotypy in the SK&F
38393-injected rats (unpublished data).
Therefore, the stereotypy-producing dopamine
D2receptors might not only cooperate with the
dopamine D1receptors but they might also be less
sensitive than the yawning-producing dopamine
D2-receptors.
The yawning produced by SND 919 or
talipexole was inhibited by the selective
dopamine D2-receptor antagonists, spiperone at a
dose of 0.5 mg/kg, YM-09151-2 at a dose of 0.1
mg/kg or the muscarinic receptor antagonist,
scopolamine at a dose of 0.5 mg/kg. However, the
yawning was unaffected by the selective dopamine
D i-receptor antagonist, SCH 23390 at a dose of
0.5 mg/kg. The above mentioned receptor
antagonist dosages were selected according to
the other experiments. It is likely that these
receptor antagonists at such doses produced
inhibitory effects on the yawning behavior
induced by SND 919 or talipexole in a
pharmacologically relevant manner and did not
exert their antagonistic actions through general
motor debilitation. Taken together, it is
assumed that the yawning observed following the
administration of SND 919, talipexole or
(+)-3-PPP is mediated by the stimulation of
specific dopamine D2-receptors, which have a
high affinity for dopamine receptor agonists
similar to that of dopamine D2-autoreceptors and
pituitary lactotroph dopamine D2-receptors.
It is well known that the tuberoinfundibular
dopamine neurons, which originate in the arcuate
and periventricular nuclei of the hypothalamus
and terminate in the external layer of the
median eminence, exert a inhibitory influence on
the release of prolactin. Moreover, dopamine D1
and D2-receptors are documented for their
important role in the regulation of prolactin
secretion. In fact, prolactin release was
inhibited by the dopamine D2-receptor agonist,
bromocriptine, and was contrarily facilitated by
the dopamine D1receptor agonist, SK&F 38393.
It has been reported that (+)-3PPP acts as a
preferential autoreceptor agonist at low doses,
and stimulates postsynaptic dopamine receptors
at high doses (Hjorth et al. 1981). Treatment
with (+)-3-PPP decreased basal prolactin levels
in naive rats and also decreased
hyperprolactinemia induced by reserpine or
alpha-MT). Talipexole has also been reported to
decrease basal prolactin levels and
reserpineevoked hyperprolactinemia. In the
present experiments, basal prolactin levels were
decreased as a result of treatment with SND 919,
talipexole or (+)-3PPP. Increases in prolactin
levels induced by alpha-MT were also antagonized
by these compounds in doses which induced a
yawning behavior. Moreover, SND 919 and
talipexole inhibited prolactin release in vitro
in slices of the rat anterior pituitary gland.
Therefore, it seems reasonable to conclude that
SND 919 can decrease prolactin secretion via
stimulating dopamine D2-receptors in the
anterior pituitary gland.
The present findings are in accord with
previous reports which show that the yawning
behavior elicited by dopamine receptor agonists
seems to concomitantly involve dopamine
D2-receptor stimulation and muscarinic receptor
activation and that prolactin release is
inhibited by dopamine D2-receptor agonists.
Furthermore, the present results suggest that
SND 919 as well as talipexole has a selective
agonistic activity for specific dopamine
D2-receptors, which are not related to the
occurrence of stereotypy and have a high
affinity for dopamine receptor agonists similar
to that of dopamine D2-autoreceptors and
pituitary lactotroph dopamine D2-receptors.