Dynamic dopamine
receptor interactions in the core and shell of
nucleus accumbens differentially coordinate the
expression of unconditioned motor
behaviors
Canales JJ, Iversen SD
Department of Experimental
Psychology, University of Oxford,
England
INTRODUCTION
The nucleus accumbens (Acb) of the ventral
striatum is a prominent structure of the basal
forebrain. Several lines of evidence support the
distinction in the Acb between a medial portion,
the shell, and a lateral sector, the core. The
shell of the Acb is connected with the lateral
hypothalamus and the neurotensin-rich
ventromedial and ventrolateral portions of the
ventral pallidum, which are regarded as
transitional areas between the basal ganglia and
the "extended amygdala". In turn, thecore of Acb
is affiliated with the motor centers of the
basal ganglia, including the dorsolateral part
of the ventral pallidum, the substantia nigra,
and the pedunculopontine nucleus. Functional and
behavioral evidence suggests that the core of
Acb is involved in motor integrative functions
typical of the basal ganglia, while the shell of
Acb mediates various aspects of incentivedriven
behavior, including feeding and
stimulant-induced reinforcement . These
functions are believed to be mediated in part by
the mesolimbic dopamine system that projects to
the core and the shell of Acb.
Dopamine exerts physiological effects in the
brain through actions on multiple dopamine
receptors. These have been grouped into two
families, the dopamine D1-class, which includes
Dl and D5 receptors and the D2-class, which
includes D2, D3, and D4 receptors. Many
behavioral effects mediated by dopamine D2-class
receptor agonists, including hyperlocomotion and
stereotypies, are dependent on simultaneous
dopamine D1class receptor activation. The
striatum is a critical site for the functional
interaction between dopamine D1-class and
D2-class receptors. For example, the
coadministration of dopamine D1-class and
D2-class receptor agonists into the Acb induces
a synergistic increase in locomotor activity.
Moreover, this treatment, but not each agonist
treatment given alone, induces stereotyped oral
behavior following injections into the
ventrolateral striatum. These findings are
consistent with the classical view, according to
which the postsynaptic effects of D2-class
receptor agonists require D1-class receptor
enabling.
We previously characterized the effects
elicited by infusions of the dopamine D2-class
receptor agonist quinpirole and the indirect
dopamine agonist d-amphetamine on unconditioned
motor behaviors following microinjections into
the striatum. The effects of quinpirole
infusions into the striatum, but not those
elicited by d-amphetamine, including sedation,
oral dyskinesia, and motor inhibition, showed a
marked homogeneity across distinct striatal
sectors. These findings prompted us to examine
whether the behavioral effects of D2-class
receptor agonists are homogenous within the Acb,
and whether the expression of specific D2-class
receptor-mediated effects following
administration into the core and the shell of
Acb depends on the level of activation of
Dlclass receptors. As a D2-class receptor
agonist we selected (+)-PD 128907, which
displays some selectivity for the dopamine D3
receptor in binding studies and functional
tests. As a D1-class receptor agonist we chose
SKF 38393, which induces potent D1-class
receptor-mediated effects in vivo despite its
partial agonist profile. These experiments were
designed to characterize the unconditioned motor
effects evoked by microinjections of (+)-PD
128907 into the core and the shell of Acb, and
to determine whether these effects, and those
previously reported for quinpirol, can be
modified by concurrent activation of the
dopamine D1-class receptor.
DISCUSSION
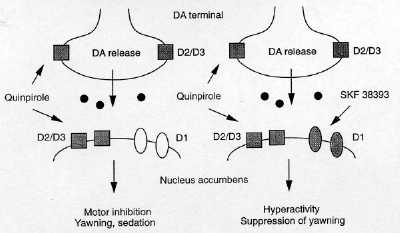
SKF 38393 switches the pattern of
responses induced by quinpirole
The dose of SKF 38393 selected in Experiment
1 was behaviorally inert, producing only
marginal effects on locomotion and oral
behaviors. Administration of this dose of SIT
38393 followed by quinpirole into the Acb
induced a striking "behavioral switch." We have
shown that animals treated with quinpirole into
the Acb display a syndrome of hypoactivity,
sedation, and oral dyskinesia. SIT 38393
virtually eliminated all behavioral signs
associated with the quinpirole syndrome.
Moreover, animals receiving SKF 38393 followed
by quinpirole into the Acb were hyperactive.
Interactions between dopamine Dlclass and
D2-class receptors have been demonstrated
previously for the induction of locomotor
activity and motor stereotypies following
systemic drug administration and intracerebral
microinjection into the Acb. We further
characterized these interactions. SKF 38393
followed by quinpirole reduced spontaneous
grooming and yawning responses, effects
typically observed following amphetamine
infusions into the Acb. Further, SIT 38393
canceled the increase in oral dyskinesia
typically induced by quinpirole injections into
the Acb. This finding suggests that abnormal
orofacial movements may result from an imbalance
in the activity of dopamine D1-class and
D2-class receptor-mediated neuronal output
pathways.
(+)-PD 128907 induces a quinpirole-like
pattern of behavioral responses
Microinjections of (+)-PD 128907 into the
core or the shell of Acb produced a strong
suppression of rearing, locomotor activity, and
grooming behavior, together with an increase in
oral dyskinesia, yawning, and sedation. This
behavioral profile is remarkably similar to that
observed after quinpirole injections into the
Acb (Canales and Iversen, 1998). (+ )-PD 128907
was equipotent at these two anatomical sites.
These results indicate that dopamine D2-class
receptors in the shell and core of Acb have
inhibitory effects on exploratory activity,
including rearing and locomotor behavior.
Systemic administration of (+)-PD 128907 and of
other agonists with limited selectivity for
dopamine D3 receptors reduces locomotor activity
in a dose-dependent fashion. Moreover, in this
study, injections of (+)-PD 128907 into the core
or shell of Acb induced intense bouts of oral
dyskinesia. Anatomical and behavioral studies
support an involvement of both core and shell
sectors of Acb in oral behaviors. While the
shell of Acb is mainly associated with the
lateral hypothalamus and the extended amygdala,
the core of Acb maintains strong links with the
substantia nigra. The accumbens-hypothalamic
pathway is known to be critical for the control
of ingestive behavior, whereas the substantia
nigra is implicated in the expression of oral
stereotypies and oral dyskinesias. (+)-PD 128907
also induced yawning and sedation in a
dose-dependent manner. At low systemic doses,
dopamine D2-class receptor agonists, e.g.,
apomorphine, quinpirole, and (+)-PD 128907, are
known to elicit sedation and yawning behavior.
Taken together, our results indicate the core
and the shell of Acb are homogeneous for the
expression of D2-class receptor-mediated
behavioral responses such as exploratory
behavior, oral dyskinesia, sedation, and
yawning.
SKF 38393 switches the pattern of
responses induced by (+)-PD 128907
Coadministration of SIT 38393 and (+)-PD
128907 into the core or the shell of Acb
revealed striking differences between these two
sites at the functional level. SKF 38393 almost
completely abolished the syndrome elicited by (+
)-PD 128907 following administration into the
core, but not into the shell, of Acb. Following
treatment with SKF 38393 and (+)-PD 128907 into
the shell of Acb, oral dyskinesia, yawning, and
sedation were significantly increased relative
to controls, while animals injected into the
core of Acb did not differ from controls in
these measures. Furthermore, a specific
synergistic interaction was observed between SKF
38393 and (+)-PD 128907 for the induction of
locomotor activity. The coadministration of SKF
38393 and (+ )-PD 128907 into the core of Acb
significantly increased locomotor activity,
while levels were significantly depressed
following administration into the shell of Acb,
compared to control levels. Thus, the
cooperative interaction between dopamine
receptors to evoke hypermotility was specific to
the core of Acb. Taken together, these results
indicate that the functional coupling between
dopamine D1-class and D2class receptors is not
homogenous within the Acb.
CONCLUSIONS
These results suggest that doses of
quinpirole and of (+ )-PD 128907 that elicit
behavioral effects classically associated with
dopamine autoreceptor activation, such as
locomotor depression and yawning, possess
"latent" postsynaptic activity at dopamine
D2-class receptors, readily uncovered by SKF
38393. A similar profile was previously
described for other D2-class receptor agonists
following systemic drug administration. The
results further suggest that, at the behavioral
level, the postsynaptic effects of quinpirole
and of (+ )-PD 128907 in the Acb depend
critically on the level of activation of
dopamine Dlclass receptors. We show that there
are two aspects to the postsynaptic effects of
D2-class receptor agonists: I behavioral
suppression and sedation following removal of
D1-class receptor tone, and behavioral arousal
and j hyperactivity following D1-class receptor
activation. Thus, dopamine D1-class receptors in
the Acb not simply enable certain dopamine
D2-class receptor-mediated behavioral responses,
but in fact regulate the switch between states
of sedation and motor inhibition to states of
strong behavioral stimulation.
A major finding in this study was that the
effects of coadministration of SKF 38393 and
(+)-PD 128907 into distinct sectors of the Acb
showed marked anatomical selectivity, suggesting
a differential sensitivity of the ( core and
shell of Acb with regards to the functional
interaction between dopamine receptors.
Morphological evidence indicated that
tyrosine-hydroxylase-contaming fibers contact
the dendrites of spiny projections neurons of
the core of Acb more frequently than those of
the shell region, and that neurons of the core
sector branch more often, have more spines and a
larger area available for synaptic interactions.
Anatomical evidence indicates that the core of
Acb preferentially interacts with motor output
structures of the basal ganglia. At the
behavioral level, the present results suggest
that there is clear functional segregation
within the ventral striaturn such that the core
of Acb exerts predominant control over the
expression of dopamine receptor mediated
unconditioned motor behaviors.
-Collins
C, Truccone A et al. Pro-erectile Effects of
Dopamine D2-like Agonists are Mediated by the D3
Receptor in Rats and Mice. J J Pharmacol Exp
Ther 2009
-Canales JJ,
Iversen SD Psychomotor activating effects
mediated by dopamine D2 and D3 receptors in the
nucleus accumbens Pharmacol Biochem Behav 2000;
67;161-168
-Canales
JJ, Iversen SD Dynamic dopamine receptor
interactions in the core and shell of nucleus
accumbens differentially coordinate the
expression of unconditioned motor behaviors
Synapse 2000; 36; 297-306
-Collins GT, JM
Witkin et al Dopamine agonist-induced
yawning in rats: a dopamine d3 receptor mediated
behavior. J Pharmacol Exp Ther
2005;314(1):310-319
-Collins GT, Newman
AH,Woods JH et al.Yawning and hypothermia in
rats: effects of dopamine D3 and D2 agonists and
antagonists. Psychopharmacology
(Berl).
2007;193(2):159-170
-Collins GT. et
al. Food restriction alters
pramipexole-induced yawning, hypothermia, and
locomotor activity in rats: Evidence for
sensitization of dopamine D2 receptor-mediated
effects. JEPT 2008;325:691-697
-Collins
GT et al. Narrowing in on compulsions:
dopamine receptor functions Exp Clin
Psychopharmacol 2008,16(4):498,502
-Chen J, Gregory T.
Collins, et al. Design, Synthesis, and
Evaluation of Potent and Selective Ligands for
the Dopamine 3 (D3) Receptor with a Novel in
Vivo Behavioral Profile. J Med. Chem.,
2008;51(19):5905&endash;5908
-Collins GT,
Truong YN, et al. Behavioral sensitization
to cocaine in rats: evidence for temporal
differences in dopamine D(3) and D (2) receptor
sensitivity. Psychopharmacology (Berl).
2011;215(4):609-20.
Li SM,
Collins GT et al. Yawning and locomotor
behavior induced by dopamine receptor agonists
in mice and rat. Behav Pharmacol. 2010; 21(3):
171&endash;181.
Based on studies in the
rat, Sokoloff et al. have made the valuable
suggestion that the D3 receptor is a
particularly important target for antipsychotics
in the mesolimbic DA system. These study in the
human demonstrates that the distribution of D3
receptors and D3 mRNA-bearing neurons is
consistent with relative segregation of the D3
subtype to the limbic striatum as well as it
primary and secondary targets and many sources
of its afferents.