Psychomotor
activating effects mediated by dopamine D2 and
D3 receptors in the nucleus
accumbens
JJ Canales, SD Iversen
Department of Experimental
Psychology, University of Oxford, UK
Introduction The mesolimbic and
nigrostriatal dopamine systems and their target
receptors play a significant role in the
expression of a wide variety of motor behaviors.
Two families of dopamine receptors have been
differentiated; the D1class, which includes the
D1 and D5 receptors, and the D2 class, which
includes the D2, D3, and D4 receptors. Members
of the family of D2 receptors share a high
degree of sequence homology and similar
pharmacological profile. Of these, one of the
most studied in recent years has been the D3
receptor. Localization studies suggest that the
expression of D3 receptors is prominent in
limbic-based circuits that modulate affect and
motivated behavior, including the islands of
Calleja and the nucleus accumbens (Acb). Given
that the expression of the dopamine D3 receptor
in the limbic system is high, this receptor site
has been postulated a a potential target for
therapeutic intervention in schizophrenia and
substance abuse.
Pharmacological evidence suggests that
dopamine D3 receptors exert inhibitory actions
on psychomotor functions. (+)-PD 128907,
pramipexole, and 7-OH-DPAT, compounds with
relative selectivity for dopamine D3 receptors.
reduce locomotor activity and induce sedation
and yawning over a wide dose range. Conversely,
D3 receptor-preferring antagonists stimulate
locomotor behavior, but while D2
receptor-preferring antagonists inhibit motor
activity. However, pharmacological studies are
not supported by behavioral studies on mice
lacking D3 receptors. The behavioral effects
induced by D3 receptor agonists, including
decreased locomotor activity and hypothermia,
are identical in wild-type and D3
receptor-mutant mice. At the functional level,
therefore, studies on the functions of the D3
receptor have been inconclusive.
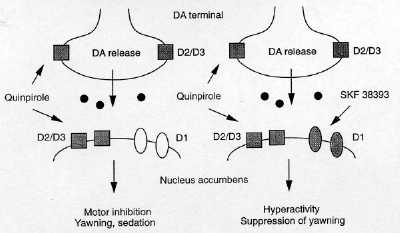
We have previously characterized the
behavioral effects of quinpirole and (+)-PD
128907, drugs with some selectivity for the D3
receptor, following injections into the Acb and
we have shown that the effects of these agonists
critically depend on the level of activation of
D1-class receptors. In the present experiments,
we first admnistered the ergot compound
bromocriptine into the Acb to compare its
behavioral effects to those induced by
microinjections of quinpirole and (+)-PD 128907.
Second, we tested whether the behavioral effects
of bromocriptine can he modified by concurrent
administration of a D1 class receptor agonist.
Third, we examined the relativ contributions of
D2 and D3 receptors to the locomotor enhancing
effects induced by coadministration of D1 class
and D2 class receptor agonists into the Acb. In
this case, we administered antagonist drugs with
relative selectivity for D2 or D3 receptors
prior to cocktail injections of D1 class and D2
class receptor agonists into the Acb. The
results are discussed in terms of the relative
contribution of D2 and D3 receptors to motor
behavior. [...]
Discussion
Typical and atypical behavioral profiles
induced bv bromocriptine injections into the
Acb: dependence on D2 class receptor
activation
Bromocriptine exhibits the typical
behavioral profile of a D2 class receptor
agonist following systemic admnistration.
Bromocriptine suppresses spontaneous locomotion
at low doses, but it induces hypermotility and
stereotyped behaviors at high doses. The
expression of some bromocriptine induced
behavioral effects depends on D1 class receptor
activation. In rats exposed to a
methyl-p-tyrosine plus reserpine, the
locomotorenhancing effects of bromocriptine are
not evident, but these can be reinstated by
concurrent administration of behaviorally
inactive doses of the dopamine D1 class receptor
agonist SKF 38393 . Studies on the effects of
bromocriptine following injections into the Acb
have produced conflicting results. Bromocriptine
decreased rat locomotor behavior in the open
field and inhibited mouse spontaneous climbing
behavior following direct injections into the
Acb. In activity cages, however, bromocriptine
induced no changes in locomotor activity. Using
a similar measure of locomotor activity and a
wider dose range, the present experiments
confirm the findings of Jenkins and Jackson and
extend their observations to other
activityrelated measures, including sniffing and
rearing. In this regard, considering the ability
of quinpirole and (+) - PD 128907 to suppress
locomotor activity following injections into the
Acb, bromocriptine is an atypical D2 class
receptor agonist.
Bromocriptine induces sedation and yawning
responses following administration of low
systemic doses. In the present experiments,
however, bromocryptine injections into the Acb
failed to elicit yawning or sedation at any of
the doses tested. In addition, bromocriptyne
produced only a weak tendency to increase oral
activity. In this respect, bromocriptine also
shows an atypical profile, because these
behaviors are elicited by injections of
quinpirole and (-I-)PD 128907 into the Acb.
These atypical effects of bromocriptine could be
due to its complex pharmacological profile.
Binding studies have shown that bromocriptine
has a high affinity for noradrenergic a1 and a2
receptors, and for, serotonin a receptors.
Moreover, microdialysis studies have shown that
bromocriptine increases 5-HT turnover, and
reduces extracellular levels of acetylcholine
and dopamine in the striatum. Thus, interactions
with neurotransmitter systems other than the
dopamine system may contribute to the atypical
D2 class receptor agonist profile of
bromocriptine following intracerebral
injections.
Previous studies suggest that D1 class
receptors may play a permissive role in the
locomotor-stimulant effects of bromocriptine
following systemic administration. In our study,
the combination of SKF 38393 and bromocriptine
enhanced locomotor activity and suppressed
spontaneous yawning responses. These two effects
are also observed following amphetamine
treatment or coadministration of SU 38393 and
quinpirole into the Acb. The present experiments
indicate that activation of dopamine D1 class
receptors at the level of Acb alters the effects
of bromocriptine in a way such that the
behavioral output clearly reflects class/D2
class receptor interactions. With regard to
these interactions, bromocriptine behaves as a
typical D2 class receptor agonist.
Studies with nafadotride and L-741626
suggest a contribution of D2 and D3 receptors in
the Acb to the induction of enhanced locomotor
behavior.
To study whether D2 and/or D3 receptors play
a role in the enhanced locomotor stimulation
induced by combinations of SKF 38393 and either
bromocriptine ór (+)-PD 128907, the
antagonist drugs L-741626 and nafadotride were
administered before injections of the agonist
drugs. The results showed that the locomotor
response elicited by SKF 38393 plus (+)-PD
128907 was effectively blocked by nafadotride at
doses that do not produce functional blockade of
D2 receptors, while L-741626 was less effective.
These results suggest that D3 receptors
synergize with D1 -class receptors and
contribute to the expression of behavioral
hyperactivity. Nafadotride also attenuated the
hyperactivity induced by SKF 38393 plus
bromocriptine, although not significantly so. In
the present experimental conditions, nafadotride
was without effect on locomotor activity.
Previously, however, stimulatory effects of
nafadotride on locomotor activity have been
observed in well-habituated rats, although this
finding has not been replicated. In the present
experiments, we did not detect any stimulatory
effects of nafadotride on locomotor
behavior.
At the doses tested, L-741 626 attenuated
the responses to (+)-PD 128907 and bromocriptine
following coadministration with SKF 38393, but
the magnitude of this attenuation did not reach
statistical significance. Considering the
overall results obtained with the antagonist
L-741626, it seems that slightly higher doses
would have blocked the locomotor effects of both
SKF 38393 plus bromocriptine and SKF 38393 plus
(+)-PD 128907. In fact, previous studies have
shown that some physiological effects of (+)-PD
128907 are readily blocked by L-741626
[6,9], suggesting that (+)-PD 128907
could lack selectivity for the D3 receptor in
vivo. Moreover, (+)-PD 128907 elicits locomotor
suppression and hypotherm both in wild-type and
D3-mutant mice, but not in D2 knock-outs. This
evidence should be carefully considered when
reaching conclusions regarding the behavioral
functions of the D3 receptor. In our study, low
doses of nafadotride significantly attenuated
the locomotor effects of the combination of SKF
38393 and (+) - PD 128907, suggesting that both
D2 and D3 receptors in the Acb sergize with D1
class receptors for the induction of enhanced
locomotor stimulation. This evidence, however,
should be evaluated with a wider range of more
potent and selective agonists and antagonists
for D3, relative to D2, receptors.
Conclusions
The results of this study indicate that
bromocriptine induces atypical D2-class
receptor- mediated behavioral effects following
administration into the Acb. However, in
combination with a D1 class receptor agonist,
bromocriptine displays the typical functional
profile of a D2 class receptor agonist. The
present observations further indicate that
antagonists with relative selectivity for D2 or
D3 receptors are able to attenuate the motor
effects induced by coadministration of a D1
class receptor agonist and either bromocriptine
or (+) - PD 128907 into the Acb. These results
suggest that in the presence of sufficient D1
class receptor activation, wich critically
modulates D2 class receptor-mediated behavioral
responses at the level of the Acb, D2 and D3
receptors in the Acb may contribute in similar
ways to the expression of heightened psychomotor
arousal.
-Collins
C, Truccone A et al. Pro-erectile Effects of
Dopamine D2-like Agonists are Mediated by the D3
Receptor in Rats and Mice. J J Pharmacol Exp
Ther 2009
-Canales JJ,
Iversen SD Psychomotor activating effects
mediated by dopamine D2 and D3 receptors in the
nucleus accumbens Pharmacol Biochem Behav 2000;
67;161-168
-Canales
JJ, Iversen SD Dynamic dopamine receptor
interactions in the core and shell of nucleus
accumbens differentially coordinate the
expression of unconditioned motor behaviors
Synapse 2000; 36; 297-306
-Collins GT, JM
Witkin et al Dopamine agonist-induced
yawning in rats: a dopamine d3 receptor mediated
behavior. J Pharmacol Exp Ther
2005;314(1):310-319
-Collins GT, Newman
AH,Woods JH et al.Yawning and hypothermia in
rats: effects of dopamine D3 and D2 agonists and
antagonists. Psychopharmacology
(Berl).
2007;193(2):159-170
-Collins GT. et
al. Food restriction alters
pramipexole-induced yawning, hypothermia, and
locomotor activity in rats: Evidence for
sensitization of dopamine D2 receptor-mediated
effects. JEPT 2008;325:691-697
-Collins
GT et al. Narrowing in on compulsions:
dopamine receptor functions Exp Clin
Psychopharmacol 2008,16(4):498,502
-Chen J, Gregory T.
Collins, et al. Design, Synthesis, and
Evaluation of Potent and Selective Ligands for
the Dopamine 3 (D3) Receptor with a Novel in
Vivo Behavioral Profile. J Med. Chem.,
2008;51(19):5905&endash;5908
-Collins GT,
Truong YN, et al. Behavioral sensitization
to cocaine in rats: evidence for temporal
differences in dopamine D(3) and D (2) receptor
sensitivity. Psychopharmacology (Berl).
2011;215(4):609-20.
Li SM,
Collins GT et al. Yawning and locomotor
behavior induced by dopamine receptor agonists
in mice and rat. Behav Pharmacol. 2010; 21(3):
171&endash;181.
Based on studies in the
rat, Sokoloff et al. have made the valuable
suggestion that the D3 receptor is a
particularly important target for antipsychotics
in the mesolimbic DA system. These study in the
human demonstrates that the distribution of D3
receptors and D3 mRNA-bearing neurons is
consistent with relative segregation of the D3
subtype to the limbic striatum as well as it
primary and secondary targets and many sources
of its afferents.