The
paraventricular nucleus of the hypothalamus
(PVN) is considered a sort of integration centre
between the central and the peripheral
autonomous nervous system and is involved in the
control of numerous functions, including male
erectile function and sexual behaviour.
Accordingly, bilateral electrolytic lesions of
the PVN reduce drug- and oxytocin-induced
erections, pheromone-mediated noncontact
erections, seen in male rats put in the presence
of an inaccessible receptive female, and impair
copulatory behaviour, while its electrical
stimulation induces erection. Several lines of
evidence show that a group of oxytocinergic
neurons originating in the PVN and projecting to
extra-hypothalamic brain areas, such as the
hippocampus, the medulla oblongata and the
spinal cord play an important role in the
control of erectile function and copulatory
behaviour. When activated, these neurons
facilitate erection and sexual activity, while a
reduced erectile function and sexual activity is
found when these neurons are inhibited. Among
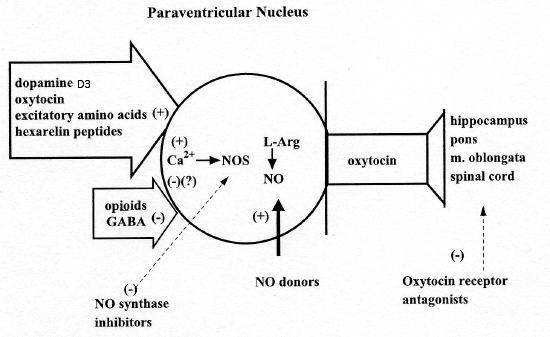
neurotransmitters and neuropeptides present in
the PVN, those that activate oxytocinergic
neurons and facilitate erection and sexual
activity include dopamine, excitatory amino
acids, and oxytocin itself, while GABA and
opoioid peptides inhibit activity of these
neurons and reduce penile erection and
copulatory behaviour. Apparently, dopamine,
excitatory amino acids and oxytocin activate
pro-erectile oxytocinergic neurons by
stimulating nitric oxide (NO) synthase, the
Ca2-calmodulin enzyme which converts L-argimne
to NO, and which is present in high
concentrations in the PVN, including the cell
bodies of oxytocinergic neurons. In line with
this hypothesis, microdialysis studies have
shown that dopamine receptor agonists,
excitatory amino acids and oxytocin itself
increase NO production in the PVN, when given at
doses that induce penile erection; the latter
effect is prevented not only by doses of NO
synthase inhibitors that also inhibit the sexual
response, but also by the opiate morphine, by
GABAA receptor agonists given into the PVN and
by oxytocin receptor antagonists. NO production
also increases in the PVN during
pheromone-induced noncontact erections and
during copulation, both these physiological
sexual response being reduced by NO-synthase
inhibitors given in the PVN, by morphine, GABAA
receptors agonists and by oxytocin receptor
antagonists. In line with the above data, the
oxytocin messenger RNA and NO synthase messenger
RNA have been found to be lower and the opioid
peptide messenger RNA higher in the PVN of
impotent male rats, when compared to sexually
potent male rats.
Although the above findings support the idea
that paraventricular oxytocinergic neurons
projecting to extra-hypothalamic brain areas and
to the spinal cord are activated during erection
and copulation, paraventricular
neurotransmitters and/or neuropeptides that
activate these neurons in physiological contexts
are still unknown. Our previous attempts to
identify these neurotransmitters and/or
neuropeptides, have revealed that noncontact
erections are partially reduced by dizolcipine
(MK-801), a potent noncompetitive excitatory
amino acid receptor antagonist of the
N-methyl-D-aspartic acid (NMDA) receptor ubtype,
while dopamine receptor antagonists and
d(CH2)5Tyr(Me))rn8-vasotocin, a potent oxytocin
receptor antagonist, were ineffective. This led
us to suggest that an increase in excitatory
amino acidergic neurotransmission may be, at
least partially, responsible for the activation
of oxytocinergic neurons mediating erection and
copulation.
However, the above results do not allow us
to rule out a possible role of paraventricular
dopamine in noncontact erections and copulation,
as the failure of dopamine receptor antagonists
to prevent the former sponses might be secondary
to the experimental conditions used. Moreover,
dopamine D2 receptor agonists are extremely
potent in inducing erection when injected into
the PVN of male rats and induce erection in both
numerous laboratory animals and in humans. In
order to provide direct evidence for a role of
dopamine in the activation of paraventricular
oxytocinergic neurons during sexual activity,
the concentration of dopamine and its metabolite
3,4-dihydroxyphenylacetic Acid (DOPAC), which
often reflects the amount of dopamine released
tnd recaptured by dopaminergic synapses and
dendrites, was measured in the paraventricular
dialysate of male rats exposed to a receptive
female both before and during copulation by
means of a highly sensitive high pressure liquid
chromatography (HPLC) method coupled to
electrochemical detection.
Discussion
To our knowledge, this is the first report
in which the concentrations of dopamine and its
main metabolite DOPAC were measured in the
dialysate obtained from male rats implanted with
vertical microdialysis probes aimed at the PVN.
Basal dopamine concentration was found to be
approximately 0.05 nM, while that of DOPAC was
approximately 0.4 nxl. As the recovery of
authentic dopamine passed throughout the probes
was found to be approximately 20%, dopamine
concentration in the paraventricular
extracellular fluid may be estimated to be close
to 0.25 nM. This value is similar to that found
in the dialysate obtained from the medial
preoptic area of male rats .
The present results show that an increase in
the concentration of dopamine and, to a lesser
extent, of its metabolite DOPAC, occurs in the
paraventricular dialysate of sexually potent
male rats, which show noncontact erections when
put in the presence of an inaccessible receptive
female rat and which copulate with the female
when permitted. The increase in dopamine and
DOPAC found during copulation was higher than
that found when copulation was not allowed. The
increase of dopamine and DOPAC concentrations
was not observed when male rats were placed with
a nonreceptive female. To our knowledge, these
findings provide the first evidence that
dopaminergic neurotransmission increases in the
PVN when penile erection occurs in physiological
contexts, such as during noncontact erections
and, to an even higher extent, during
copulation, when 'in copula' erections occur.
Indeed, the increase of dopamine parallel to
that of DOPAC in the paraventricular dialysate
of male rats, reasonably reflects the activation
of paraventricular dopaminergic neurons during
sexual activity. In this regard it is pertinent
to recall that dopamine in the PVN is present in
neurons and synapses that belong to
incertohypothalamic dopaminergic neurons. These
neurons have their cell bodies in the A13 and
A14 dopaminergic groups of Dalhstrome and Fuxe,
arborize extensively and impinge on neurons in
several hypothalamic nuclei, including the PVN
and the medial preoptic area.
These findings are in line with previous
studies showing that dopamine D2 receptor
agonists injected into the PVN induce penile
erection, facilitate penile reflexes and
influence copulation. As recalled in the
Introduction, dopamine receptor agonists
facilitate erectile function and copulatory
activity when injected into the PVN by
activating oxytocinergic neurons projecting to
extra-hypothalamic brain areas, such as the
hippocampus, and the spinal cord.
Accordingly, doses of apomorphine that
induce erection, increase oxytocin content in
the hippocainpus and in blood, and
apomorphineinduced erections are abolished by
bilateral electrolytic lesions of the PVN, which
deplete oxytocm across the central nervous
system or by oxytocin receptor antagonists given
centrally, with a potency that is parallel to
the potency of these compounds in blocking
oxytocmergic receptors. Oxytocin receptor
antagonists are also effective in reducing
noncontact erections and copulatory behaviour of
sexually potent male rats, as well as the
facilitory effect of apomorphine on copulatory
behaviour in sexually potent male rats.
The mechanism by which dopamine activates
oxytocinergic neurons through the stimulation of
D2 receptors located in the cell bodies of
oxytocinergic neurons mediating erectile
function is apparently mediated by an increased
Ca 2+ influx inside the cell bodies of these
neurons. This causes in turn the activation of
NO-synthase, thereby increasing NO production in
the PVN. NO in turn activates oxytocinergic
neurons mediating erectile function by an as yet
unidentified mechanism. Accordingly,
apomorphine-induced penile erections, noncontact
erections and copulation occur concomitantly
with an increased NO production in the PVN, as
measured by the increase in concentration of NO2
and NO3, the main metabolites of newly formed
NO, in the dialysate obtained from the PVN
during these sexual responses. Furthermore, the
latter responses are all strongly reduced by NO
synthase inhibitors injected into the PVN, which
also reduce NO production in the PVN.
Conversely, the injection of classic NO donors
into the PVN induces erections indistinguishable
from those induced by drugs (apomorphine,
oxytocin and NMDA). Although numerous
experimental data support the above hypothesis,
other mechanisms of the action of dopamine in
the facilitation of penile erection and sexual
activity at the PVN level cannot be ruled out.
As already extensively discussed, dopamine may
also activate oxytocinergic neurons mediating
erectile function by removing an inhibitory
input on them other than by acting directly on
D2 receptors located in their cell bodies.
However, irrespective of D2 receptor location in
the PVN, and of the fact that in many tissues D2
receptors inhibit Ca 2+ influx through
voltage-dependent Ca 2+ channels, nanogram
amounts of the potent N-type voltagedependent Ca
2+ channel inhibitor, w-conotoxin, injected into
the PVN, prevent penile erection induced by
apomorphine. This finding is in line with the
hypothesis that an increased Ca 2+ influx into
the cell bodies of oxytocinergic neurons plays a
key role in the activation of these neurons and
of penile erection by dopamine and its
agonists.
The increase in the concentration of
dopamine and DOPAC that occurs in the
paraventricular dialysate of sexually potent
male rats put in the presence of a receptive
female, and to an even higher extent during
copulation, resembles the increase of the
concentration of dopamine and DOPAC that occurs
in the dialysate obtained from the medial
preoptic area of sexually potent male rats in
similar experimental conditions. Also in these
studies dopamine and DOPAC concentrations
increased in the dialysate from the medial
preoptic area only when sexually potent male
rats were usd with a receptive female, while no
increase was found when copulati ri did not
occur or when castrated male rats were used.
Another area in which microdialysis studies
provided evidence of an increase in dopamine
concentration during sexual activity, is the
nucleus accumbens, which contains the nerve
endings of mesolimbic dopaminergic neurons that
play a key role in sexual motivation and
rewarding. Together these findings suggest that
dopamine in the PVN may play a role not only in
erectile function (e.g. a component of the
consumatory phase of sexuaI behaviour) but also
in sexual motivation, as suggested for the
medial preoptic area. However further studies
are necessary to verify such a possibility.
Recently, we found that the mixed D1-D2
receptor antagonist cisflupentixol, the
selective D1 receptor antagonist SCH 23390, and
the selective D2 receptor antagonist raclopride,
injected into the PVN were unable to prevent
noncontact erections, as was the oxytocin
receptor antagonist
d(CH2)5-T'r(Me)Orn8-vasotocin, while th NMTDA
receptor antagonist dizolcipine (MK-801) was
able to reduce, although only partially, this
sexual response. Apparently, these findings seem
to support the hypothesis that dopamine in the
PVN is not involved in pheromone-mediated
noncontact erections. This contrasts with the
findings of the present study which show that
dopamine is increased in the paraventricular
dialysate of male rats that show this sexual
response when put in the presence of an
inaccessible receptive female. Indeed, one would
speculate that if dopamine activity were
increased in the PVN during noncontact
erections, these would have been found to be
reduced by the prior administration of dopamine
receptor antagonists into the PVN. However, in
the light of the present results, this is
unlikely. In fact, the inability of dopamine
receptor antagonists to reduce noncontact
erection might have been due to reasons other
than a lack of a role for dopamine in this
sexual response. For instance, the doses of
dopamine receptor antagonists used in the
previous study may have been too low or the
volume injected not sufficient to spread and act
across the entire PVN, as all dopamine
antagonists were injected unilaterally.
Alternatively, and most likely, the PVN might be
a sort of node of parallel circuits that mediate
either the precopulatory or the copulatory phase
of sexual behaviour, so that all parallel
circuits would have to be blocked in order to
prevent this sexual response.
In conclusion, the present study further
confirms that the PVN plays a primary role in
the control of male rat sexual behaviour. Our
results show for the first time that dopamine
neurotransmission increases in this hypothalamic
nucleus not only when sexual response is induced
by drugs and/or neuropeptides, but also when it
occurs in physiological contexts, such as
noncontact erections or during copulation. In
line with previous studies, it is likely that
dopamine activates its own receptors, increasing
in turn NO production inside paraventricular
oxytocinergic cell bodies in the PVN, thereby
activating oxytocinergic neurons projecting to
extra-hypothalamic brain areas and in the spinal
cord, which control erectile function and sexual
behaviour. The PVN might be one of the sites in
which dopamine receptor agonists (e.g.
apomorphine) act to facilitate erectile function
not only in rats but also in humans.