Department of Physiology,
Toho University School of Medicine, Ohta-ku,
Tokyo. Japan
-Kita
I, Kubota N, Yanagita S, Motoki C
Intracerebroventricular administration of
corticotropin-releasing factor antagonist
attenuates arousal response accompanied by
yawning behavior in rats. Neurosci.Letter
2008;433(3):205-208
-Kita
I, Yoshida Y, Nishino S. An activation of
parvocellular oxytocinergic neurons in the
paraventricular nucleus in oxytocin-induced
yawning and penile erection. Neurosci Res.
2006;54(4):269-275
-Kita I,
Sato-Suzuki et al.Yawning responses induced
by local hypoxia in the paraventricular nucleus
of the rat.Behavioural Brain Research
2000;117(1-2):119-126
-Kubota
N, Amemiya S, Motoki C, Otsuka T, Nishijima T,
Kita I. Corticotropin-releasing factor
antagonist reduces activation of noradrenalin
and serotonin neurons in the locus coeruleus and
dorsal raphe in the arousal response accompanied
by yawning behavior in rats. Neurosci Res.
2012;72(4):316-323
-Seki Y, Y
Nakatani, et al Light induces cortical
activation and yawning in rat Behav Brain Res
2003;140(1-2):65-73
-Seki Y,
Sato-Suzuki I, et al Yawning/cortical
activation induced by microinjection of
histamine into the paraventricular nucleus of
the rat. Behav Brain Res.
2002;134(1-2):75-82.
-Sato-Suzuki I,
Kita I, Oguri M, Arita H Stereotyped yawning
responses induced by electrical and chemical
stimulation of paraventricular nucleus of the
rat Journal of Neurophysiology,
1998;80(5)2765-2775
Abstract : We examined the effects of
light stimulation on cortical activation and
yawning response in anesthetized, spontaneously
breathing rats. Cortical activation was assessed
by means of an electrocorticogram (ECoG) and
yawning response was evaluated by monitoring an
intercostal electromyogram as an index of
inspiratory activity and a digastric
electromyogram as an indicator of mouth opening.
Light stimulation elicited an arousal shift in
the ECoG to faster rhythms. This arousal
response was followed by a single large
inspiration with mouth opening, i.e. a yawning
response. Higher light intensity significantly
reduced the onset latency of the arousal/yawning
response. Pretreatment with pyrilamine, an
H1-histamine receptor antagonist, injected into
the lateral ventricle blocked both the cortical
activation and the yawning response induced by
light stimulation, suggesting a role of brain
histaminergic neurotransmission in modulating
the light-induced arousal yawning
responses.
1. Introduction : Yawning is a very
common behavior in humans and other animals, yet
it has received little attention in science as
well as in daily life. The lack of interest in
yawning behavior may be due to the lack of
knowledge of its physiological significance. One
approach to clarify the functions of yawning is
to reliably evoke a yawning response and monitor
the response together with the concomitantly
occurring associated phenomena. In this
connection, we have recently reported that a
stereotyped yawning response can be evoked by
chemical stimulation of the paraventricular
nucleus (PVN) of the hypothalamus in
anesthetized, spontaneously breathing rats. In
those studies, we recorded the
electrocorticogram (ECoG) to evaluate arousal
responses during yawning, and found that ECoG
arousal, represented by lower voltage and faster
rhythm, occurred before the yawning behavior,
which indicates that yawning has physiological
significance in increasing alerting
mechanism.
Although the PVN is known to be essential
for the occurrence of yawning, our previous
studies raised another fascinating possibility:
that the PVN may also play an important role in
triggering cortical activation. We have shown
that the PVN mediates the arousal yawning
response induced by higher brain ischemia. It
was also suggested that the PVN plays an
important role in the arousal yawning responses
evoked by orexinergic
as well as histaminergic
neurotransmission. Since the PVN is generally
known to play a critical role in stress
responses, the PVN may also be involved in
cortical activation related to stress.
In this study we sought to determine whether
bright light stimulation, one kind of acute
stress, can induce cortical activation together
with the yawning response. This idea was first
suggested by two lines of evidence. The first is
that bright light has immediate alerting effects
in humans as well as rats. The second is that
yawning, a behavior generally believed to be
induced by drowsiness or boredom, is also
influenced by the circadian
rhythm in humans as well as rats.
Everybody might have yawned due to light
stimulation when waking up in the morning.
Indeed, it is reported that yawning is most
likely in the morning shortly after waking, a
condition corresponding to a transition from
dark to light, but the neural mechanisms
underlying the light-induced arousal effect or
yawning are poorly understood. The present study
was designed to explore whether the PVN is
involved in this mechanism.
By using our model of the arousal yawning
response, we examined whether brief light
stimulation of the eyes of anesthetized,
spontaneously breathing rats induces the
cortical activation as well as the yawning
response. Attempts were also made to identify
whether histamine, a neurotransmitter considered
to be important in regulation of the arousal
system, yawning and the circadian rhythm, is
involved in these responses. [...]
4. Discussion : This is the first
study showing the effects of light stimulation
on arousal yawning response in anesthetized
rats. Light stimulation induced yawning,
together with an arousal shift in the ECoG;
Since yawning is a behavior mediating the PVN of
the hypothalamus, we speculate that the PVN
might be also involved in the light-induced
arousal signaling pathway.
The concept that the PVN is essential for
the occurrence of yawning was first proposed by
the Argiolas and Melis group, who found that
microinjection of several substances, including
apomorphine, into the PVN increases the
frequency of spontaneous yawns in freely moving
rats. Consequently, they demonstrated that
lesions of the PVN prevent yawning induced by
apomorphine. On the other hand, we provided
evidence that a stereotyped yawning response can
be evoked by several chemical stimulations of
the PVN in anesthetized, spontaneously breathing
rats. The yawning response has important
physiological significance in anesthetized
animals, since various physiological aspects
accompanied to the yawning behavior, such as the
autonomic or arousal responses can be
concomitantly monitored. We further found that
it is the medial
parvocellular subdivision(mp) of the
PVNwhich is involved in the yawning response.
Within the mp, the neurons: responsible for
yawning might be the oxytocinergic:
parvocellular neurons projecting to the lower
brain stem. This suggestion was principally
based on the report of Sawchenko and Swanson who
demonstrated that oxytocinergic parvocellular
neurons in the PVN send descending axons to the
lower brain stem, a region involved in arousal,
respiratory, cardiovascular and other autonomic
functions. In view of this, we suggest the
possibility that light stimulation induces the
yawning response by indirectly activating the
oxytocinergic parvocellular neurons in the PVN
projecting to the lower brain stem. However, we
cannot exclude the possibility that other
pathways, besides the oxytocinergic ones, can be
involved in the yawning response. For instance,
ACTH injected into the PVN and surrounding
periventricular region induces yawning, that is
not involving oxytocinergic pathways.
What kind of signaling pathway is then
involved in the light-induced yawning response
mediated by the PVN? Light is the predominant
stimulus in maintaining the circadian rhythm in
humans as well as other animals. The principal
pacemaker that generates circadian rhythms is
located in the suprachiasmatic nucleus (SCN) of
the hypothalamus, and the light signal reaches
the SCN through the retina-hypothalamic tract
from the retina. Among the various target areas
of the SCN, the parvocellular part of the PVN is
a region especially important because it plays a
role in autonomic functions that are influenced
by circadian rhythins. For example, light
stimulation modifies the autonomic nervous
system in rats via the SCN and via further
projection to the parvocellular part of the PVN.
As mentioned above, the parvocellular part of
the PVN is also the site responsible for the
yawning response, a behavior known to be
influenced by the circadian rhythm. Therefore,
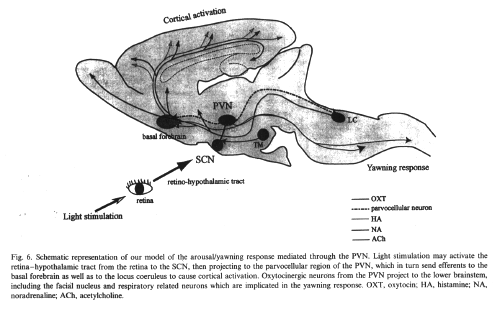
the light signal may first be transmitted from
the retina to the SCN, which in turn projects to
the PVN, then mediating the yawning
response.
Our hypothesis that the light-induced
yawning response is mediated by signals conveyed
from the retina to the SCN and further to the
PVN is also supported by several pieces of
histological evidence. Daikoku et al. reported
that in rats light exposure induces a remarkable
enhancement of c-fos immunoreactivity in neurons
within the SCN and the parvocellular part of the
PVN. More evidence comes from the observation
that light induces c-fos expression in the SCN
output neurons targeting the PVN [. Although
not directly to the stress-related area of the
PVN, Buijs et al. showed projections from the
SCN to the periventricular and rostral PVN
together with the dorsomedial hypothalamus, the
regions known to project into the PVN, and
proposed a mechanism that input from the SCN to
the PVN could be influenced by either stress or
environmental factors, such as light.
Although we suggest that the signals from
the retina to the SCN and further projection to
the PVN is responsible for the light-induced
yawning response, we cannot exclude the
possibility that other inputs to the PVN may
mediate this response. For instance, signals
conveyed from the retina to the superior
colliculuspretectum, another region known to be
involved in light perception, could be related
to this response, but until now there has been
no report showing direct projection from the
superior colliculus-pretectum to the PVN.
As to neurotransmission from the SCN to the
PVN, several transmitters have been reported.
The most essential one is the vasoactive
intestinal polypeptide (VIP) of the SCN which
plays a role in the control of the autonomie
nervous system. Subsequently, VIP as well as
gastrin-releasing peptide was shown to be
involved in the light-activated output neurons
of the SCN. On the other hand, glutamate and
GABA were shown to mediate rapid
neurotransmission from the SCN to the
parvocellular region of the PVN in rats.
Considering our previous data showing that
microinjection Of L-glutamate induces an arousal
yawning response, L-glutamate might be one
neurotransmitter mediating the light-induced
response in the present study. Nevertheless, the
possibility of other transmitters, especially
VIP, being involved in this response still needs
clarification.
We demonstrated that light stimulation
induces the yawning response together with an
arousal shift in the ECoG, which suggests that
the PVN is involved not only in the
light-induced yawning response but also in the
light-induced cortical activation. The role of
the PVN in arousal regulation has been
consistently reported in our previous studies.
For example, the PVN mediates the arousal
pathway induced by higher brain ischemia. It is
also suggested that the PVN plays an important
role in the arousal responses evoked by
orexinergic as well as histaminergic
neurotransmissien. We have suggested that the
projection from the PVN to the basal forebrain
or the locus coeruleus could account for the
cortical activation. In all of these studies, a
yawning behavior accompanied the arousal
response, which indicates that the PVN mediates
the cortical activation related to the yawning
behavior. In view of these notions, the
light-induced arousal response observed in the
present study may be mediated by inputs from the
retina to the SCN, then projecting to the
parvocellular region of the PVN, which in turn
sends efferents to the basal forebrain as well
as the locus coeruleus to cause cortical
activation.
Since the PVN is a region implicated in
stress responses, it may also be involved in
cortical activation induced by various
stressors. In this regard, the bright light used
in the present study can be regarded as a kind
of novel stress. Although not directly related,
it should be noted that anatomical projections
of the SCN to stress-related areas of the PVN
have been reported and connections between the
SCN and PVN are affected by stress.
A unique aspect of the yawning response is
that a depressor response always precedes the
final yawning event (a single large inspiratory
effort) induced by light stimulation. The
reduction in autonomie responses by light
stimulation is supported by recent data of
Scheer et al. who demonstrated a reduction in HR
after light exposure in rats. One may raise a
question whether the depressor response observed
in the present study by light stimulation might
be an arousal response or rather the reverse as
could be inferred from the fact that light in
nocturnal rodents suppresses activity. In this
concern, we observed an arousal shift in the
ECoG concurrent with the depressor response in
the present study. In addition, we reported in
our previous studies that the depressor/arousal
response occurs not only by light stimulation
but also by local hypoxia of the PVN as well as
the yawning responses induced by L-glutamate,
nitric oxide donor or orexin. As we mentioned
above, the yawning response together with the
depressor response may be mediated by the
oxytocinergic PVN neurons descending to the
lower brain stem. On the other hand, the arousal
response induced by light stimulation may be
mediated by ascending pathway from the PVN to
areas responsible for cortical activation.
Corticotropin-releasing factor (CRF) neurons
within the PVN could be nominated for such
ascending pathway, however this is not yet clear
and we are currently examining this
possibility.
Histamine is a neurotransmitter involved in
yawning as well as in the arousal system. It has
also been suggested that histamine participates
in the regulation of the circadian rhythm.
Although histamine-containing neuronal cell
bodies are restricted to the hypothalamic
tuberomammillary nucleus (TM), there is a high
concentration of histamine in the SCN. Indeed
anatomically, fibers are known to arise in the
TM and project to the SCN. These facts give rise
to the suggestion that histamine participates in
the regulation of the circadian rhythm. In this
study we examined whether histamine could modify
the light-induced arousal yawning response.
Pretreatment with HI-histamine receptor
antagonist in the lateral ventricle blocked the
light-induced yawning as well as the cortical
activation, indicating that these responses are
modified by histamine neurotransmission. It has
been shown that histamine has excitatory and
inhibitory effects on neurons in the SCN.
Whereas the excitatory effect of histamine is
mediated by the HI receptor in the SCN, the
inhibitory effect is mediated by the H2
receptor. Therefore, the neurons responsible for
the light-induced arousal yawning response
within the SCN could have been blocked by HI
antagonist in the present study.
Administration of HI antagonist into the
lateral ventricle caused a shift to slower waves
in the ECoG which is consistent with the studies
using mepyramine, another HI-histamine receptor
antagonist, causing an increase in cortical slow
waves. These data are also consistent with the
sedation and drowsiness caused in man by
antihistaminics, together confirming the role of
histaminergic neurotransmission in arousal
regulation.
In conclusion, light stimulation elicited a
yawning response together with cortical
activation. Since yawning is a behavior mediated
through the PVN, the results suggest that the
light-induced yawning response as well as the
arousal response is mediated by signals from the
retina to the SCN and further projection to the
PVN. These findings further strengthen our
hypothesis that the PVN plays a significant role
in the arousal mechanism related to,
yawning.
-Kita
I, Kubota N, Yanagita S, Motoki C
Intracerebroventricular administration of
corticotropin-releasing factor antagonist
attenuates arousal response accompanied by
yawning behavior in rats. Neurosci. Lettre
2008;
-Kita
I, Yoshida Y, Nishino S. An activation of
parvocellular oxytocinergic neurons in the
paraventricular nucleus in oxytocin-induced
yawning and penile erection. Neurosci Res.
2006;54(4):269-275
-Kita I,
Sato-Suzuki et al.Yawning responses induced
by local hypoxia in the paraventricular nucleus
of the rat.Beh Brain Res 2000; 117; 1-2; 119 -
126
-Sato-Suzuki I,
Kita I; Oguri M, Arita H Stereotyped yawning
responses induced by electrical and chemical
stimulation of paraventricular nucleus of the
rat Journal of Neurophysiology, 1998; 80, 5;
2765-2775
-Seki Y, Y
Nakatani, et al Light induces cortical
activation and yawning in rat Behav Brain Res
2003; 140; 1-2; 65-73
-Seki Y,
Sato-Suzuki I, et al Yawning/cortical
activation induced by microinjection of
histamine into the paraventricular nucleus of
the rat. Behav Brain Res.
2002;134(1-2):75-82.
-Collins
G, JM Witkin et al Dopamine agonist-induced
yawning in rats: a dopamine d3 receptor mediated
behavior J Pharmacol Exp Ther 2005
-Hipolide DC; Lobo
LL; De Medeiros R; Neumann B; Tufik S
Treatment with dexamethasone alters yawning
behavior induced by cholinergic but not
dopaminergic agonist. Physiol Behav 1999; 65;
4-5; 829-32
-Hipolide
DC, Tufik S Paradoxical sleep deprivation in
female rats alters drug-induced behaviors
Physiol Behav. 1995; 57; 6; 1139-1143
-Moyaho A,
Valencia J Grooming and yawning trace
adjustment to unfamiliar environments in
laboratory Sprague-Dawley rats J Comp Psychol
2002; 116; 3; 263-269
-Neumann BG,
Troncone LR, Braz S, Tufik S Modifications
on dopaminergic and cholinergic systems induced
by the water tank technique: analysis through
yawning behavior. Arch Int Pharmacodyn Ther
1990; 308; 32-38
-Tufik S et
al Effects of stress on drug induced yawning
Physiol Behav 1995; 58; 1; 1881-1884