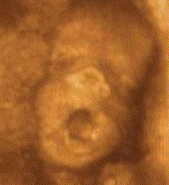
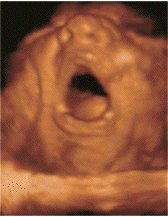
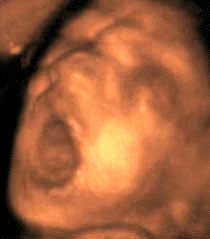
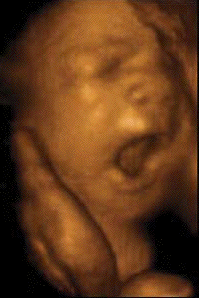
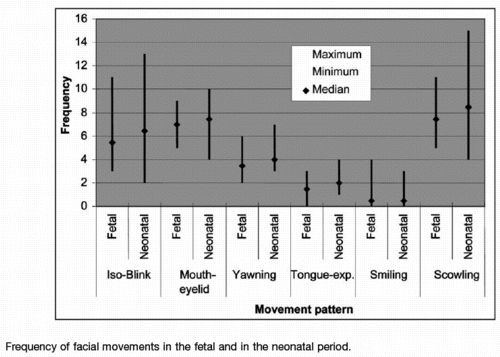
Sleep associated with a high frequency of phasic
activities, particularly muscle twitches, dominate the daily
life of altricial mammals and human neonates.
Piontelli
A. Fetal Yawning in Development of
Normal Fetal Movements. The fisrt 25 weeks of
gestation. Spinger. 2010.
135p.
Résumé: L'échographie
4D permet l'évaluation des expressions
faciales du foetus et en particulier de
reconnaître un comportement banal, le
bâillement. Bien qu'il s'agisse d'un
comportement pluri-quotidien, le
bâillement a suscité peu
d'intérêt tant en recherche qu'en
pratique médicale. Phylogenèse et
ontogenèse permettent de proposer une
théorie de son origine. Comme un
interrupteur de type "va et vient", l'alternance
d'action des structures cérébrales
stimulant l'éveil ou le sommeil engendre
l'émergence de différents
états de vigilance. Succédant
à l'hypotonie musculaire du sommeil
paradoxal avec lequel le bâillement
partage des liens ontogénétiques,
l'étirement musculaire puissant qu'il
représente active par
rétro-contrôle les structures du
tronc cérébral impliquées
dans l'éveil. L'existence du
bâillement chez le foetus témoigne
d'un développement harmonieux du tronc
cérébral, de la fonction
neuro-musculaire périphérique et
de l'installation de rythmes ultradiens de la
vigilance. La reconnaissance de l'absence de
bâillements comme de mouvements de
déglutition, associée ou non
à un rétrognatisme prédit
un risque de dysfonctionnement postnatal du
tronc cérébral. (in
english)
La pratique de l'échographie foetale
permet de remarquer quotidiennement un
comportement: le bâillement. Peu de
données sont parues dans la
littérature concernant le
bâillement foetal. Les auteurs y
révèlent leur perplexité:
"le bâillement est universel mais mal
compris" (1) ou "un réflexe rudimentaire
n'ayant qu'une fonction obscure, s'il en a une"
(2). Bien que bâiller soit un acte
quotidien peu de recherches lui ont
été consacré. Nous nous
proposons de lui donner sens et d'indiquer en
quoi il peut enrichir l'exploration
échographique. En préambule, il
est nécessaire de préciser que la
recherche, chez l'homme, sur les
développements comportementaux foetaux
n'est jamais directement opératoire mais
le plus fréquemment basée sur des
homologies avec d'autres
vertébrés.
Un dicton populaire prédit que
"l'organe crée la fonction". Pourtant,
l'embryologie nous enseigne que les mouvements
du foetus sont nécessaires à la
maturation neuro-fonctionnelle de la
motricité et participent au
développement d'autres organes comme les
poumons. L'activité motrice du foetus
indique également l'installation d'une
organisation fonctionnelle harmonieuse tant du
système nerveux central que de la
fonction neuro-musculaire
périphérique (3).
Tous les mouvements qu'un nouveau-né
est capable d'exécuter ont
été rodés au cours de vie
foetale et perdurent toute l'existence. Les
comportements observés au cours de la vie
intra-utérine comme la respiration, le
bâillement et d'autres ont un continuum
avec ceux observés après la
naissance et participent de la
neurogénèse anatomo-fonctionnelle
(4).
L'apparition et l'évolution
ontogénétique de la
motricité foetale sont
étudiées depuis l'existence de
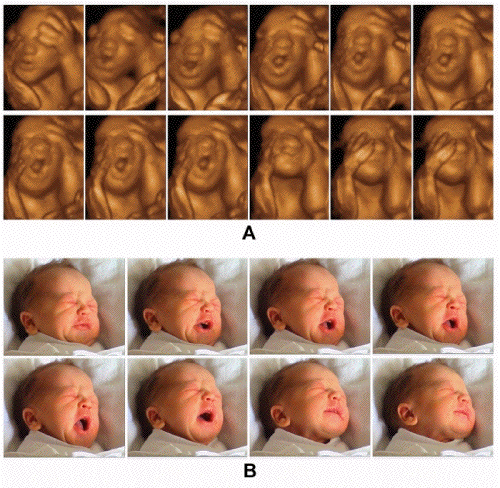
l'échographie. L'échographie en 3D
animée ou 4D perfectionne cet apport en
autorisant l'évaluation non seulement de
mouvements mais de comportements
spécifiques. La distinction des
expressions de type émotionnel du visage
n'était pas possible avec
l'échographie 2D ou 3D. C'est grâce
à l'échographie 4D qu'il est
possible de préciser le sourire et aussi
le déroulement harmonieux des
différentes composantes du
bâillement (5,6).
Ce que le bâillement n'est
pas
R.R. Provine et B.C. Tate (7) ont
infirmé l'hypothèse, largement
répandue, d'une amélioration de
l'oxygénation cérébrale par
le bâillement. Ni
l'élévation du CO2 ni la baisse de
l'O2 sanguin ne déclenche de
bâillements. Des sujets respirant de
l'oxygène pure bâillent et la
fréquence de leurs bâillements
n'est pas réduite. L'existence de
bâillements foetaux, en milieu liquidien
amniotique, comme les bâillements des
poissons, confirment l'incapacité du
bâillement à améliorer
l'oxygénation
cérébrale.
L'atélectasie, c'est à dire le
collapsus des alvéoles, résulte
d'une hypoventilation alvéolaire.
L'activité ventilatoire normale de
l'adulte comporte de soupirs et d'amples
inspirations (le bâillement en est une)
qui préviennent l'atélectasie en
concourant à la dispersion du surfactant
alvéolaire. La bâillement foetal en
milieu liquidien ne peut engendrer aucune
prévention de l'atélectasie alors
que la production de surfactant n'apparaît
qu'à la naissance (8)
Comment reconnaître un
bâillement ?
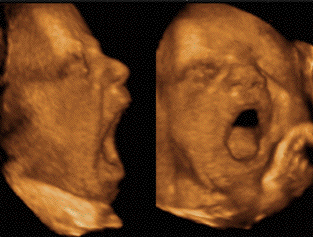
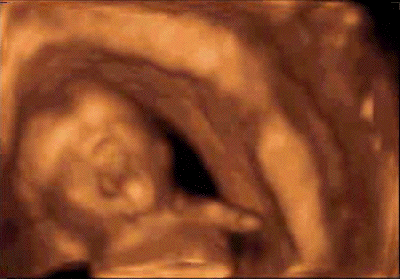
Le bâillement est un cycle
paroxystique comportant en 5 à 10
secondes une succession de mouvements, toujours
dans la même chronologie:
-une ample, lente et profonde inspiration,
avec une bouche grande ouverte. Chez l'adulte
l'expansion du pharyngo-larynx quadruple son
diamètre de repos.
- une brève apnée à
thorax plein, fréquemment
accompagnées de mouvements
d'étirement des membres et du tronc
(pandiculation)
-une rapide expiration passive (9)
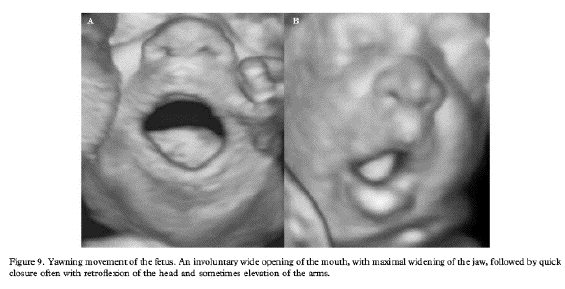
L'écho 4D permet de
différentier ces différents temps.
La bouche du foetus, précédemment
close, s'ouvre au maximum pendant 4 à 6
secondes avec une rétraction de la
langue, suivie d'une rapide fermeture
combinée avec une dorsi-flexion de la
nuque et parfois élévation des
bras derrière la tête
(pandiculation). Cette harmonieuse
séquence motrice est nettement
différente d'une rapide
déglutition (10). En usant du doppler
couleur, il est possible de visualiser un flux
liquidien amniotique pénétrant
dans la bouche, l'oropharynx, la trachée
jusqu'aux poumons. Cette séquence reste
isolée, non répétée,
contrairement aux bâillements de l'adulte
survenant fréquemment en
série.
Le bâillement n'est pas une simple
ouverture de bouche mais correspond à un
étirement massif des muscles
respiratoires (le diaphragme et les
intercostaux), des muscles de la face et du cou.
Cette association synergique de mouvements
complexes réalise un comportement
très stéréotypé
qu'on peut qualifier de réflexe en raison
de son déclenchement involontaire. L'arc
réflexe parcourt l'hypothalamus, la
substance réticulée du tronc
cérébral, les noyaux du complexe
pré-Bötzinger de la ventilation, les
noyaux moteurs des nerfs crâniens (V,VI,
IX,X, XI, XII) et cervicaux commandant le
diaphragme et les muscles intercostaux (C1-C4).
Ainsi il peut être inféré
que le bâillement est une part d'un
étirement généralisé
des muscles anti-gravitaires qui l'accompagne
fréquemment, la pandiculation
(9).
Embryologie et mécanismes
En 1973, T. Dobzhansky écrivait "rien
n'a de sens, en biologie, sauf à le
regarder sous l'angle de l'Evolution"(12). E.
Haeckel (1834-1919) déclarait
"l'ontogenèse est une brève et
rapide récapitulation de la
phylogenèse, déterminée par
les lois de l'hérédité
(transmission génétique,
inné) et de l'adaptation
(phénotype, acquis)" (13). Le
bâillement illustre parfaitement ces
sentences. D'une part, l'échographie en
confirme la précocité
ontogénétique, puisqu'il
apparaît entre 12 et 15 semaines de la vie
intra-utérine (4). D'autre part, le
bâillement est un comportement
phylogénétiquement ancien,
retrouvé des reptiles aux
mammifères, chez les
vertébrés des mondes terrestres,
aériens et sous marins. Sa survivance,
sans variation évolutive, postule de son
importance tant d'un point de vue fonctionnel
que développemental. La puissante
contraction musculaire qu'il représente a
un coût métabolique
élevé. Si nous nous accordons aux
principes des lois de l Evolution
établies par Ch. Darwin, ce coût
doit être contrebalancé par un
avantage évolutif et de
développement. Ainsi une hypothèse
structurale postule un accroissement
d'activation et de recrutement des
neurotrophines qui génèrent toute
une cascade de nouvelles synapses, de nouveaux
circuits neuronaux au niveau
diencéphalique et du tronc
cérébral. Ce mécanisme de
développement,
activité-dépendant, a clairement
été identifié comme un des
processus affectant la maturation précoce
neuro-comportementale des systèmes
sensoriels et moteurs. Ce
phénomène
d'activité-dépendance est
probablement un processus ubiquitaire de
maturation cérébrale par lequel le
développement d'une région, d'une
structure participe au développement
d'autres régions, d'autres structures
(14).
La capacité à
générer un comportement moteur
à commande centrale et à le relier
à l'éveil et à la
respiration est une propriété de
la formation réticulée du tronc
cérébral qui est,
phylogénétiquement,
remarquablement conservée au travers des
poissons, des amphibiens, des reptiles et des
oiseaux. Il en découle que la
conservation des mécanismes
développementaux orchestrant
l'organogenèse du tronc
cérébral chez tous les
vertébrés est probablement crucial
pour l'éveil et la respiration (15).
Un grand nombre de données ont
été collectées sur les
gênes exprimés chez l'embryon et
qui gouvernent la segmentation de la tête,
du cou et du système nerveux. Les
gènes Hox représentent quatre
groupes de gênes codant pour la
transcription de facteurs impliqués dans
l'orchestration de la mise en place de l'axe
rostro-caudal de l'organisme, incluant la
segmentation du tronc cérébral et
du diencéphale d'une part de la formation
des membres supérieurs d'autre part (15).
C'est là, probablement, l'explication de
l'origine des anomalies de développement
cranio-facial que l'échographie aide
à dépister et de la sentence qui
énonce "la face prédit le
cerveau".
Le massif facial et le cerveau
s'individualisent à partir d'une
structure embryonnaire commune, l'ectoblaste. Le
pôle céphalique comporte une
segmentation originelle embryologique
encéphalo-faciale et
encéphalo-cervicale avec une
correspondance topographique stricte: les
structures naso-frontales et
prémaxillaires sont liées au
cerveau antérieur; les structures
maxillo-mandibulaires et cervicales
antérieures sont unies au tronc
cérébral et à ses nerfs. Au
début du troisième mois, l'embryon
devient un fœtus grâce à
l'apparition des premières
séquences motrices orales et
pharyngées sous la dépendance du
développement neurologique du tronc
cérébral: développement de
l'activité de succion-déglutition
et de bâillement. Succion et
bâillement ont donc la même origine
embryologique, soulignant l'importance du tronc
cérébral dans le
développement neuro-physiologique de
l'activité oro-pharyngée
coordonnée avec les régulations
respiratoire, cardiaque et digestive de
même localisation neuro-anatomique.
L'apparition de ces comportements, vers douze
semaines de grossesse, permet de dater le stade
où le tronc cérébral est
individualisé et l'hypophyse devient
fonctionnelle, alors que l'extension du
néocortex temporal et frontal se
complète jusqu'à 22 à 24
semaines (17,18).
Les mouvements des joues et de la langue
participent à la formation du palais par
l'initiation de mouvements de traction
antéro-postérieurs alors que les
valves palatales primordiales sont
orientées verticalement.
L'activité motrice de la langue et du cou
sont constamment accompagnés de
mouvements de la bouche (19). Les liens entre
les structures neurales commandant la
motricité oro-linguale et les centres
coordonnant respiration et motricité des
bras ne sont pas parfaitement compris. Il semble
que les informations sur les rythmes
respiratoires et moteur des membres,
nécessaires au cervelet afin de les
coordonner, convergent vers le noyau
réticulaire latéral du tronc
cérébral (20).
Le bâillement résulte de
l'activation de multiples neurotransmetteurs et
neuropetides au niveau du système nerveux
central. Le noyau paraventriculaire de
l'hypothalamus est le centre hypothalamique qui
adapte et coordonne les réponses
hormonales et autonomiques caractérisant
ce comportement. Ses neurones ocytocinergiques
sont stimulés par la dopamine, les acides
aminés excitateurs,
l'acétylcholine, la sérotonine,
l'oxyde nitrique, l'ACTH et les neuropeptides
apparentés (tous impliqués dans
l'éveil) alors que les opioïdes sont
inhibiteurs. Ils projettent vers des aires
cérébrales extra-hypothalamiques
telles l'hippocampe et le tronc
cérébral qui jouent un rôle
central dans l'expression motrice du
bâillement. D'autres neurotransmetteurs,
tels la noradrénaline,
l'hypocrétine, et les hormones sexuelles
influencent le bâillement, non dans son
déroulement, mais dans son
déclenchement et sa fréquence
(21).
Pourquoi le bâillement est-il
relié à l'éveil ?
La phylogenèse suggère que le
repos nocturne des poïkilothermes a
probablement évolué vers le
sommeil paradoxal (REM sleep) qui est
caractérisé par une hypotonie
musculaire périphérique
commandée par des noyaux situés
partie dorsale du tronc cérébral,
situé rostralement par rapport au pont
(22).
A partir de nombreuses études ayant
concerné tant le foetus humains que
l'animal, les données recueillies
aboutissent à considérer que la
première forme de sommeil a des
caractéristiques de sommeil actif ou
agité qui représente une forme
immature de sommeil paradoxal et est encore
très prépondérant à
la naissance. Il en découle que le
sommeil calme, forme immature du sommeil
profond, émerge peu à peu,
à mesure que le temps en sommeil
paradoxal se réduit pendant toute la
période d'ontogenèse (23).
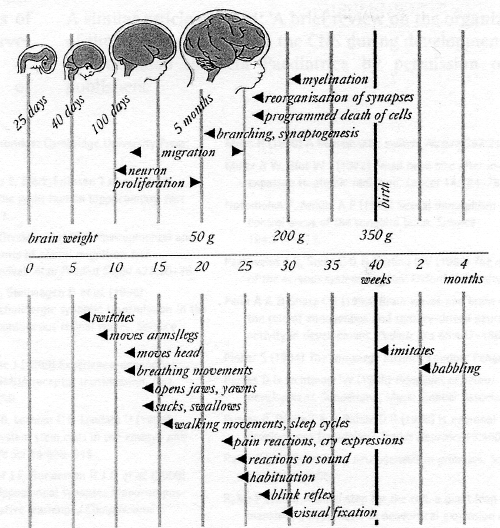
Au cours la vie intra-utérine, le
foetus présente, initialement, une
activité motrice
désordonnée, phasique et cyclique
qui se coordonne peu à peu. Il est
possible de distinguer des périodes
d'activité et de repos ou de sommeil.
ALTernent des phases de secousses myocloniques
et d'agitation motrice importante (type
locomotion, étirement, bâillement)
à tonus musculaire élevé et
des phases d'immobilité avec aspect
d'hypotonie. L'activité motrice des
membres tels les coups de pieds et les
étirements, les bâillements sont
considérés comme
caractéristiques d'une période
d'éveil. Les phases de myoclonies
succèdent à une période
d'immobilité et sont
généralement suivies de
façon brusque par une intense
activité motrice formant comme un cycle
sommeil - veille. Bien que la présence de
myoclonies soit plus importante en
intensité et en fréquence chez
l'enfant qu'en période de sommeil
paradoxal de l'adulte, leurs similarités
avec le comportement adulte et la succession
constante à l'hypotonie suggèrent
une continuité développementale
entre les phases de sommeil du foetus à
l'enfant et à l'adulte. La maturation du
système nerveux central,
caractérisée par sa
myélinisation, suit un parcours
caudo-rostral depuis la moelle puis le tronc
cérébral jusqu'aux
hémisphères. C'est pourquoi, les
mécanismes commandant le sommeil
paradoxal sont les premiers fonctionnels et les
seuls actifs initialement. Ensuite le sommeil
lent apparaît quand les structures
thalamo-corticales deviennent matures. Il
apparaît donc que le contrôle de
l'activité neuronale exercée par
le sommeil paradoxal participe du
mécanisme, activité
dépendant, de maturation fonctionnelle du
cortex. Il peut être inféré
qu'au tout début de la vie foetale, le
sommeil paradoxal (et le bâillement?)
dirige l'évolution de la maturation
corticale par sa stimulation neuronale
(24).
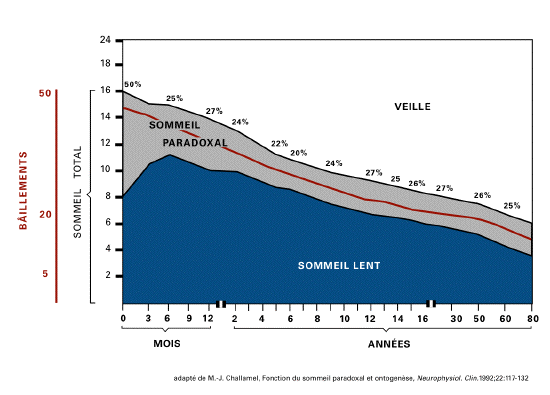
De la vie prénatale à la vie
postnatale, un pattern comportemental montre un
développement parallèle de
l'ontogenèse du sommeil paradoxal et du
bâillement. C'est ainsi que la
durée du sommeil paradoxal décline
de 50% du temps de sommeil, chez le
nouveau-né à une à deux
heures chez l'adulte, que le nombre de
bâillements passe de 30 à 50 par
jour chez le nouveau né à moins de
20 par jour chez l'adulte. Cette diminution
intervient essentiellement de la naissance
à la fin de la puberté (25).
L'apparition des différents
états de vigilance est associée
à un changement spectaculaire de leurs
durées et de leurs
périodicités. On remarque, en
premier, l'instauration d'un rythme ultradien.
Sur une période de 50 à 60 minutes
se succèdent des alternances
d'activité et d'immobilité, comme
chez le nouveau-né. La fréquence
de un à deux bâillements à
l'heure peut être associée à
ce rythme. Les bâillements s'observent
environ deux semaines avant que les rythmes de
vigilance soient discernables puis peu à
peu coalescent à leur
périodicité. Aucun changement de
la fréquence des bâillements n'a
été noté de la 20
ème à la 36 ème semaine de
grossesse par P.J. Rodenburg at al. (26). Chez
le nouveau-né à terme, des
bâillements sont très souvent
notés tout au long des premiers jours de
vie. L'embryon et le foetus sont exposés
à un rythme circadien maternel qui peut
jouer un rôle dans le développement
des pacemakers foetaux. Aucune donnée n'a
été publiée sur la place
que le bâillement pourrait indiquer comme
marqueur de l'interaction entre les rythmes
maternels et foetaux.
C. Saper et al. (27) propose un
modèle d'interactions réciproques
entre les systèmes neuronaux
d'éveil et d'endormissement, un peu comme
un interrupteur de type va et vient. Ce
modèle permet d'appréhender la
rapide transition constatée de
l'éveil au sommeil lent profond et de la
sortie du sommeil paradoxal à
l'éveil. Cette capacité
apparaît comme une nécessité
pour la survie en autorisant, d'une part, un
repos réparateur, et d'autre part, la
fuite rapide face à un prédateur
(éveil). Cette transition est sous le
contrôle des structures autonomiques qui
agissent de façon intégrée
et avec une capacité d'anticipation. Le
bâillement peut s'interpréter comme
un comportement (une partie de la pandiculation)
témoignant de cette étape
transitionnelle entre les états de
sommeil et d'éveil, c'est à dire
un mécanisme de renforcement du tonus
musculaire. L'éveil est sous la
dépendance de quatre circuits neuronaux
redondants situés dans le pont
(adrénergique), dans le pédoncule
(dopaminerqiue), dans l'hypothalamus
(histaminergique), dans la région
basi-frontale de Meynert (cholinergique). Le
système permissif contrôlant
l'éveil doit être stimulé de
façon tonique et permanente par le
système à hypocrétines de
l'hypothalamus latéral (son
déficit est la cause de la narcolepsie).
A l'opposé, l'activation des neurones du
noyau VLPO de l'hypothalamus antérieure,
en regard du chiasma optique, induit
l'entrée en sommeil profond. La puissante
contraction musculaire que représente le
bâillement stimule par rétroaction
les zones de la réticulée du tronc
cérébral impliquées dans
l'éveil (locus coeruleus). Lors de
l'éveil bâillement et pandiculation
inversent l'hypotonie musculaire qui
caractérise le sommeil paradoxal. D'autre
part, quand la pression de sommeil augmente, il
est supposé que l'activation des neurones
gabaergique et galaninergiques du VLPO diminuent
le tonus des muscles anti-gravitaires, notamment
ceux de la nuque et des masséters que le
bâillement inverserait. F. Giganti et al.
ont constaté chez des
prématurés des bâillements
lors de tous les moments du
nycthémère à l'exception de
la période de sommeil calme (lent). Ils
interprètent les bâillements comme
un état de transition, suggérant
qu'ils aient un effet stimulant des
comportements moteurs faciaux. L'ensemble de ces
données permet de proposer que le
bâillement, à l'image d'une boucle
réflexe, agit comme renforcement des
mécanismes d'éveil lors de
périodes transitionnelles de vigilance
(28,29,30).
Bâiller ou pas : une pathologie
?
Les bâillements surviennent avec une
récurrence régulière de
l'ordre de un à deux par heure. Quand un
bâillement est constaté au cours
d'un examen échographique 4D, il s'agit
le plus souvent d'un hasard ou le
résultat d'un examen
particulièrement prolongé. Le
bâillement apparaît après un
période de repos et signifie un
éveil. Si un mouvement de
déglutition a pu être
observé (beaucoup plus fréquent),
la recherche de l'existence de bâillements
n'apporte pas de données
supplémentaires. Inversement, l'abscence
ou une dysfonction de la déglutition
encourage à prendre un temps
prolongé d'examen afin d'observer
l'ensemble de l'activité motrice du
foetus et ainsi de distinguer les
différents comportements
caractérisant son rythme ultradien.
L'absence de bâillement et de
déglutition doit conduireà
rechercher une hypoplasie mandibulaire et une
glossoptose associé avec une fente
palatine.
B. Petrikovsky et al. ont rapporté
que des salves de bâillements
étaient observées dans une
série d'anémies foetales et
indiquaient qu'une telle observation de
bâillements répétés
était une aide à la recherche de
signes d'anémie foetale.
Maladies sans données
collectées
L'enfant doit être capable de respirer
et de se nourrir afin d'assurer sa survie
après la naissance. Ceci n'est possible
que par l'activation de structures anatomiques
et d'une coordination centrale adequate qui
coordonnent la succion-déglutition, la
ventilation, le sommeil et l'éveil. Le
bâillement est associé à
chacun de ces comportements. Nous avons
essayé d'établir une liste, non
exhaustive, de cas où l'étude des
bâillements foetaux pourrait avoir un
intérêt.
Le syndrome d'Ondine ou
d'hypoventilation centrale congénitale
comporte des épisodes de pauses
ventilatoires, essentiellement nocturnes par
défaut d'automatismes bulbaires, avec une
relative insensibilité à
l'hypercapnie et à l'hypoxie, en
l'absence de toute anomalie cardio-respiratoire.
Le syndrome d'Ondine peut être
associé à la maladie
d'Hirschprung. Un défaut de la
déglutition et de la motricité
oesophagienne a été
identifié chez quelques
nouveaux-nés atteints de dysmorphies
faciales et d'hypotonie.Ceci suggère une
anomalie diffuse de la motricité
digestive et de la commande au niveau du tronc
cérébral.
L'hypoplasie mandibulaire est une
anomalie fréquente des structures
craniofaciales et peut être
différenciée en forme
congénitales et
développementales.
-dysostose mandibulofaciale avec
variétés d'anomalies des
membres
-La séquence de Pierre Robin
est caractérisée par un palais
ogival, un rétrognatisme et une
glossoptose. De nombreux arguments plaident pour
une origine embryonnaire consistant en une
anomalie du développement du tronc
cérébral. Les difficultés
d'alimentation sont constamment au premier plan
fonctionnel. Des témoignages de parents,
recueillies par l'un d'entre nous, semblent
indiquer l'absence de bâillements chez les
nouveaux-nés atteints et l'apparition en
parallèle, progressivement au cours de la
première année de vie, des
automatismes de la déglutition et des
bâillements. Ainsi le syndrome de Pierre
Robin peut s'interpréter comme une
dysfonction des automatismes engendrés
par le tronc cérébral, à
l'origine des défauts de
développement anatomique des structures
mandibulaires par déficit de stimulation
motrice coordonnée. Son dépistage
est possible dès 23 semaiens de grossesse
par une échographie 3/D/4D (33,34).
-Tout syndrome (avec anomalies de croissance
uni ou bilatérale) asoocié ou non
à une ankylose de l'articulation
temporo-mandibulaire: syndrome de Franceschetti,
syndrome de Goldenhar, syndrome de
Richner-Hanhart.
Le
syndrome de Moebius asssocie une
diplégie faciale, une paralysie
occulomotrice par involution des noyaux des
nerfs VI VII, et XII. L'association avec des
malformations des membres suggère une
interruption lors de la morphogenèse du
tronc cérébral entre la 4° et
la 7° semaine de grossesse. La paralysie du
voile du palais et d'autres structures
concourant à la déglutition sont
responsables de la dysphagie qui peut s'amender
progressivement pendant l'enfance.
L'incapacité à fermer la bouche
est la règle (35).
Les infarctus artériolaires
terminaux du tronc cérébral,
constitués pendant la vie foetale,
s'expriment par différentes neuropathies
avec déficits du contrôle central
de la respiration et de la déglutition
(36).
Le syndrome de Goldenhar comprend des
malformations, le plus souvent
hémicorporelles, touchant joue, bouche et
oreille. Il représente l'expression d'un
déficit d'embryo-formation des premier et
deuxième arcs branchiaux et touchant l'os
temporal (37).
Le syndrome de Joubert est une
désordre génétique rare
caractérisé par l'absence ou
l'hypo-développement du vermix
cérébelleux et d'autres
malformations du tronc cérébral.
Le plus fréquemment, existent une ataxie,
des déficits du contrôle de la
ventilation, des apnées du sommeil, des
déficits moteurs de la langue, des yeux,
et une hypotonie.
Le trismus congénital, le syndrome de
Crisponi, le syndrome de
Stüve-Widemann....
Conclusion
La pratique de l'échographie
prénatale est passée
progressivement d'une expertise anatomique
à une évaluation anatomique et
fonctionnelle, grâce aux perfectionnements
techniques, notamment de l'apport des
technologies 3D et 4D. La reconnaissance du
bâillement foetal peut, d'une part, aider
à appréhender l'harmonie du
développement neuro-fonctionnel du tronc
cérébral, et d'autre part,
à évaluer l'installation des
mécanismes neuronaux des systèmes
d'éveil et de sommeil. Des anomalies de
son occurrence accréditent la
nécessité d'approfondir
l'exploration afin de rechercher, en cas
d'excès, une anémie foetale, en
cas de manque, une dysharmonie évolutive
et fonctionnelle du tronc cérébral
associée ou non une hypoplasie
mandibulaire. L'intérêt pour le
bâillement, que nous souhaitons avoir
éveillé par cette publication,
doit servir de moteur pour de futures
recherches, compte-tenu du très faible
contingent de données actuellement
répertoriées dans un très
large panel de malformations foetales et de
maladies néonatales.
5. Kurjak
A, Stanojevic M, Azumendi G et al. The
potential of four dimensional (4D)
ultrasonography in the assessment of fetal
awareness. J Perint Med. 2005;33:46-53.
6.Hata
T, K Kanemshi, et al. Real-time 3-D
sonographic observation of fetal facial
expression J Obstet Gynaecol Res 2005; 31; 4;
337-340
11. Masuzaki H,
Masuzaki M. Color Doppler imaging of fetal
yawning. Ultrasound Obstet Gynecol.
1996;8(5):355-356.
12. Dobzhansky T. Nothing in biology makes
sense except in the light of evolution. The
American Biology Teacher 1973;35:125-129.
13. von
Haeckel E. Anthropogenie oder,
Entwickelungsgeschichte des menschen, Keimes-
und stammesgeschichte. Leipzig : W. Engelmann
ed. 1877: 770p.
14. Briscoe J, Wilkinson DG. Establishing
neuronal circuitry: hox genes make the
connection. Genes Dev.
2004;18(14):1643-1648.
15. Graham A. The development and evolution
of the pharyngeal arches. J Anat.
2001;199:133-141.
16. Köntges G, Lumsden A.
Rhombencephalic neural crest segmentation is
preserved throughout craniofacial ontogeny.
Development. 1996;122:3229-3242.
17. Jacob J, Gutrhie S. Facial visceromotor
neurons display specific rhombomere origin and
axon pathfinding behavior. J Neuosci.
2000;20:7664-7671.
18. Santagati F, Rijli F. Cranial neural
crest and the building of the vertebrate head.
Nature Rev Neurosci. 2003;4: 806-818.
19. Wragg LE, Smith JA, Borden CS. Myoneural
maturation and function of the fetal rat tongue
at the time of secondary plate closure. Arch
Oral Biol. 1972;17:673-682.
20. Ezure
K, Tanaka I. Convergence of central
respiratory and locomotor rhythms onto single
neurons of the lateral reticular nucleus. Exp
Brain Res. 1997;113:230-242.
22. Argiolas
A, Melis MR. The neuropharmacology of
yawning. Eur J Pharmacol. 1998;343(1):1-16.
21. Siegel JM. Sleep phylogeny : clues to
the evolution and function of sleep. In Luppi PH
ed. Sleep : circuits and functions. CRC Press
2005;9:163-176.
23. Valatx JL. The ontogeny and physiology
confirms the dual nature of sleep states. Arch
Ital Biol. 2004;142(4):569-80.
24. Blumberg MS, Luca DE. A developmental
and component analysis of active sleep. Develop
Psychobiol.1996;29(1):1-22.
25. Kobayashi T, Good C, Mamiya K, et al.
Develppment of REM sleep drive and clinicals
implications. J Appl Physiol.
2004;96:735-746.
26. Roodenburg
PJ, Wladimiroff JW, van Es A et al.
Classification and quantitative aspects of fetal
movements during the second half of normal
pregnancy. Early Hum Develop.
1991;25:19-35.
27. Saper
CB, Chou TC, Scammell TE. The sleep switch:
hypothalamic control of sleep and wakefulness.
Trends Neurosci. 2001;24(12):726-31.
29. Pace-Schott EF, Hobson A. The
neurobiology of sleep: genetics, cellular
physiology and subcortcal networks. Nature Rev
Neurosci. 2002;3(8):591-605.
30. Baenninger
R. On yawning and its f Baenninger R. On
yawning and its functions. Psychonomic Bul Rev.
1997;4(2):198-207.
31. Petrikovsky
BM, Kaplan GP, Holsten N. Fetal yawning
activity in normal and high-risk fetuses: a
preliminary observation. Ultrasound Obstet
Gynecol. 1999;13:127-130.
32. American Thoracic Society.
Idiopathic congenital central hypoventilation
syndrome: diagnosis and management. Am J Respir
Crit Care Med 1999;160:368-373.
33. Abadie
V, Morisseau-Durand M. Brainstem
dysfunction: a possible neuroembryological
pathogenesis of isolated Pierre Robin sequence.
Eur J Pediatr. 2002;161(5):275-280.
34. Matsumato M, Yanagihara T et al.
Antenatal three-dimensional sonographic features
of Pierre Robin Syndrome. Case report. Gyncol
Obstet Invest. 2001;51(2):141-142.
35. May M, Schaitkin B, Shapiro A: Facial
nerve disorders in newborns and children. In The
Facial Nerve. Thieme Medical Publishers 2000;
2nd edition:339-365.
36. Sarnat HB. Watershed infarcts in the
fetal and neonatal brainstem: an aetiology of
central hypoventilation, dysphagia, micrognatia.
Europ J Paed Neurol. 2004;8(2):71-87.
37. Volpe P, Gentile M. Three dimensional
diagnosis of Goldenhar syndrome. Ultrasound
Obstet Gyncol. 2004;24(7):798-800.
Abadie
V Développement de la face: face et
séquence de Pierre Robin Compte-rendu du
5° congrès de médecine
foetale 2000
Kim
TH, Lee JJ, et al. Efficacy of assessment in
fetal behaviour by four dimensional
ultrasonography. J Obstetrics Gynaecology
2010;30(5):439-443
Kurjak
A, Azumendi G, et al Fetal hand movements
and facial expression in normal pregnancy
studied by four-dimensional sonography J.Perinat
Med. 2003;31:496-508
Kurjak
A, Azumendi G, et al The potential of 4D
ultrasonography in the assessment of fetal
awareness J Perinat Med 2005; 33; 46-53
Kurjak
A, M Stanojevic et al Behavioral pattern
continuity from prenatal to postnatal life a
study by four dimensional (4D) ultrasonography J
Perinat Med 2004;32:346-353
Masuzaki H
Color Dopplerimaging of foetal yawning
Ultrasound in obstetric & gynecolgy
1996;8(5):355-6
Petrikovsky
B, Kaplan G, Holsten N Fetal yawning
activity in normal and high-risk fetuses: a
preliminary observation. Ultrasound Obstet
Gynecol 1999;13(2):127-130
Piontelli
A. Fetal Yawning in Development of Normal
Fetal Movements. The fisrt 25 weeks of
gestation. Spinger. 2010. 135p.
Reissland
N, Francis B, Mason J. Development of Fetal
Yawn Compared with Non-Yawn Mouth Openings from
24-36 Weeks Gestation. Plos One.
2012;7(11):e50569
Roodenburg
PJ et al Classification and quantitative
aspects of fetal movements during the second
half of normal pregnancy Early Human Development
1991;25:19-35
This Photo was taken by Wolfgang
Moroder. Feel free to use my photos, but please
mention me as the author and send me a message.
(Travail personnel) [CC-BY-SA-3.0
(http://creativecommons.org/licenses/by-sa/3.0)
ou GFDL
(http://www.gnu.org/copyleft/fdl.html)], via
Wikimedia Commons