Résumé : Certains cas
d'hémiplégies associent
l'élévation involontaire du bras
paralysé au déroulement du
bâillement, ce que nous avons
baptisé « parakinésie
brachiale oscitante ». C'est un
phénomène curieux, reconnu depuis
plus de 150 ans, apparaîssant rare parce
que, sans doute, le plus souvent
négligé. Quatre observations ont
été recueillies dans le service de
neurologie du CHU de Poitiers. Leur description
détaillée est rapportée.
Une explication physiopathologique est
avancée. La levée du
contrôle cortical permet à des
structures sous-corticales,
phylogénétiquement plus anciennes,
de retrouver une autonomie au cours d'un
processus d'homéostasie associant la
stimulation de la vigilance au contrôle de
la ventilation. Ce mouvement ventilatoire est
couplé à la motricité chez
les quadrupèdes. Ainsi ressurgit lors du
cycle ventilatoire du bâillement un
automatisme du bras, comparable au balancement
de celui-ci lors de la marche.
Abtract
: In some cases of hemiplegia the onset of
yawning is associated with an involuntary
raising of the paralysed arm. For this
phenomenon we coined the term «
parakinesia brachialis oscitans ».
This curious phenomenon, that was first
described some 150 years ago, is seldom
reported, probably because it is mostly
neglected. Four observations have been made in
the neurology unit of the (University) Hospital
of Poitiers. A detailed description of these
cases is reported and a physiopathologic
explanation is proposed. The cessation of
cortical inhibition allows some subcortical
structures, which are phylogenetically more
ancient, to regain autonomy in the homeostasis
process, which associates the stimulation of
vigilance with breathing control. In quadrupeds
breathing is linked intimately with locomotion
(walking). Thus, during the onset of yawning (as
a special type of breathing) the paralysed arm
automatically starts moving as in the balancing
act of walking.
Dans la Gazette
médicale de Strasbourg 1851, le
Dr Liégey de Rambervillers rapporte cette
observation : «Un homme, étant en
convalescence d'une fièvre intermittente
hémiplégique, fut pris d'un
bâillement très incommode se
reproduisant chaque jour à la même
heure. Chose non moins remarquable, chaque fois
que ce bâillement avait lieu, il se
produisait un mouvement convulsif
d'élévation au bras
antérieurement paralysé, mouvement
accompagné d'une sensation douloureuse
dans cette partie, et que le malade ne pouvait
empêcher qu'en saisissant fortement ce
membre avec la main du côté
opposé». Notre propos est de
présenter ici, à partir
d'observations cliniques récentes, ce
phénomène curieux, associant la
mobilisation involontaire d'un membre
supérieur paralysé à un
bâillement et de proposer une
hypothèse physiopathologique. La
première observation actuellement
documentée date de 1835.
Observation n°1 : Mr. M.
René, né le 30 novembre 1925,
droitier non fumeur, est traité pour un
diabète non insulino-dépendant et
une hypercholestérolémie, sans
hypertension artérielle. En novembre
1996, il présente un déficit
moteur isolé du membre inférieur
gauche, résolutif en 24 heures.
L'exploration de cet accident ischémique
transitoire met en évidence des
sténoses bilatérales des carotides
internes de 40 à 50 %, avec des plaques
pariétales de la bifurcation carotidienne
droite d'échostructure inhomogène,
non ulcérées, sans thrombus.
Quelques jours plus tard, l'imagerie par
résonnance magnétique (IRM)
cérébrale ne montre pas de
lésion résiduelle.
L'échographie cardiaque est normale. Il
lui est prescrit 250 mg par jour d'aspirine. En
septembre 1999, s'installe brutalement un
vertige rotatoire, rapidement suivi d'une
hémiplégie gauche, globale et
proportionnelle, sans trouble sensitif, sans
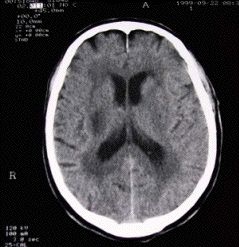
nystagmus, sans hémianopsie. La
tomodensitométrie (TDM)
cérébrale est normale à son
arrivée. Le doppler des vaisseaux du cou
apporte les mêmes données que celui
réalisé auparavant. Le holter ECG
ne retrouve ni trouble du rythme ni signe
d'ischémie. L'échographie
cardiaque trans-oesophagienne met en
évidence un thrombus intra-auriculaire
gauche. La TDM de contrôle objective
secondairement une hypodensité d'allure
ischémique du centre semi-ovale droit et
du noyau lenticulaire droit avec une
leucoaraïose périventriculaire.
Le diagnostic d'occlusion d'origine
embolique des artères
lenticulostriées, branche de la
cérébrale moyenne droite est
retenu. Le patient est mis sous Héparine,
avec relais par AVK. Un an plus tard, le patient
garde une hémiplégie gauche
sévère à
prédominance brachio-faciale; la
spasticité est importante. Lors d'une
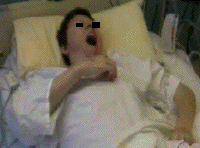
consultation de suivi, il signale une
élévation involontaire du membre
supérieur gauche, totalement
paralysé, survenant pendant le
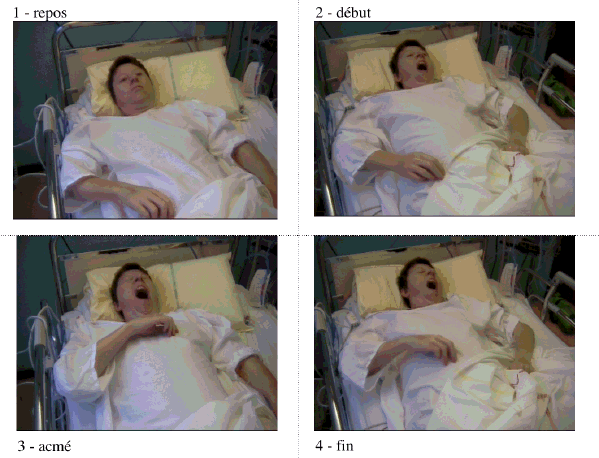
bâillement. En 2 à 3 secondes, le
bras s'élève d'une dizaine de
centimètres et se porte en adduction,
avec flexion du coude, de façon synchrone
à l'ouverture de la bouche. Il retombe
inerte à sa fermeture. Ceci peut survenir
alors qu'il est assis, debout ou couché.
Il avait constaté ce
phénomène dès le
début de son hémiplégie.
Trois ans après l'accident vasculaire, en
juin 2002, ce mouvement involontaire du bras
paralysé apparaissant lors du
bâillement persistait.
Observation n°2: Mr. B.
Paul, né le 8 avril 1946, droitier, est
porteur connu d'un déficit en
protéine C activée (mutation du
facteur V de Leiden traitée par AVK).
Cette anomalie de l'hémostase a
déjà été responsable
de phlébites du membre inférieur
droit en 1999 et 2000. En plus de ce facteur de
risque de thrombose, il a une
hypercholestérolémie à 2,4
g/l et fume environ 15 cigarettes par jour
depuis l'âge de 14 ans. Entre le 27 mai
2001 et le 02 juin 2001, il présente une
répétition d'accidents
ischémiques cérébraux
transitoires, à bascule,
caractérisés par des
déficits moteurs à
prédominance brachio-faciale, justifiant
son hospitalisation. Le doppler carotidien et
l'échocardiographie sont normaux; il a
constamment une pression artérielle
normale. Le 6 juin 2001, alors que
l'héparine est arrêtée
depuis 4 heures, afin de réaliser une
artériographie des troncs
supra-aortiques, s'installe brutalement une
hémiplégie gauche, globale et
proportionnelle, avec déficit sensitif,
sans hémianopsie et qui ne
régressera pas. La TDM met en
évidence un infarctus du noyau
caudé et du centre semi-ovale droits.
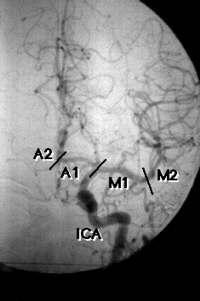
L'artériographie visualise une ectasie de
la carotide droite à type de
dolicho-artère de la portion C2 et C3 du
siphon carotidien, un thrombus de
l'artère cérébrale moyenne
à l'origine des artères
lenticulostriées. Dans les heures suivant
la constitution de l'infarctus
cérébral, Mr. Paul B., très
somnolent, bâille profondément de
façon anormalement fréquente et
répétée. Au début de
chaque bâillement, de façon
involontaire, le membre supérieur gauche
s'élève avec une flexion du coude
et adduction du bras. Ce geste dure le temps du
bâillement. La force musculaire
développée est si importante que
son épouse, assise à
côté de lui, n'arrive pas à
s'y opposer.
Observation n°3 :
Mr. L. Fernand, né le 14 septembre 1923,
droitier, cirrhotique d'origine
éthylique, présente le 7 novembre
2001, une hémiparésie droite
proportionnelle brachio-céphalique
régressant en une heure. Il a 14/8 de
pression artérielle mais, les jours
suivants, sa pression artérielle
systolique sera très variable avec
plusieurs pics supérieurs à 20.
Résidant dans un service de
gériatrie, il n'a pas de doppler mais un
scanner considéré comme normal. Il
reçoit une injection de Lovenox 40 par
jour. Le 10 novembre 2001, s'installe une
hémiplégie droite
brachio-céphalique avec parésie
linguale, une aphasie de Broca, une
hypopallesthésie au membre
supérieur. La pression artérielle
est à 22/10. Thrombopénie à
90 000. Le doppler retrouve des lésions
identiques d'aspect au niveau des carotides
internes droite et gauche, à type de
plaques pariétales calcifiées
irrégulières, de 1,5 cm de haut,
non ulcérées sans thrombus
adhérent, sténosantes à
50%, sans retentissement hémodynamique.
Les artères vertèbrales sont
normales. L'échocardiographie montre une
oreillette gauche dilatée, une
hypertrophie ventriculaire gauche concentrique,
une insuffisance mitrale modérée,
une absence de thrombus intra-auriculaire. Le
rythme cardiaque est sinusal à l'ECG,
sans trouble conductif. Le scanner
réalisé le 19 novembre (J+9)
montre un infarctus capsulo-lenticulaire gauche,
sans trace hémorragique, sans effet de
masse. L'ischémie résulte d'une
thrombose de l'artère
cérébrale moyenne au niveau des
artères lenticulostriées.
Dès l'installation de son accident, il
est noté une fréquence accrue des
bâillements. Lors de chacun, le bras droit
s'élève et se porte en adduction,
avec flexion du coude, puis retombe dès
l'occlusion buccale.
Observation n°4 : Mme. M.
Christine, née le 15 mai 1968,
droitière, fumeuse, présente au
7ème mois de sa troisième
grossesse une plébite superficielle du
mollet (juin 2003). Elle reçoit une
injection quotidienne de Lovenox 40 pendant un
mois. Le 20 octobre 2003, soit deux mois
après son accouchement, sous
contraception orale par œstroprogestatifs
minidosés, s'installe brutalement une
céphalée inhabituelle, diffuse,
intense, rebelle à tous les antalgiques
de palier I essayés. Elle ne consulte pas
de médecin. Le 22 octobre 2003, son mari
la découvre au sol, paralysée et
aphasique. A l'arrivée aux urgences, sont
notées: une hémiplégie
droite complète massive, une aphasie
totale, des mydriases bilatérales peu
réactives, une agnosie, l'absence de
raideur méningée, de syndrome
cérébelleux et de trouble
sensitif. La malade est très somnolente
et manifeste une nette dissociation
automatico-volontaire. Pression
artérielle à 12/7, rythme
cardiaque sinusal. La TDM réalisée
à l'arrivée est normal, sans
hémorragie méningée. Le
doppler est normal à droite. A gauche,
occlusion complète de la carotide interne
gauche, sans aucun flux d'aval.
L'échocardiographie trans-oesophagienne
est normale. Une TDM réalisée
à distance en novembre 2003 (J+20) montre
un infarctussylvien completet une prise de
contraste cortical
fronto-temporo-pariétale gauche.
L'artériographie retrouve une dysplasie
fibromusculaire et une dissection carotidienne
gauche, responsable d'un infarctus sylvien
cortico-sous cortical complet. Dès la
phase initiale, au stade d'hémiplagie
flasque complète, il est noté que,
lors de chaque bâillement, son bras droit
totalement paralysé,
s'élève avec flexion du coude, se
porte en adduction vers le visage puis retombe
lourdement lors de la fermeture de bouche.
L'étiologie est soit un accident
vasculaire ischémique ou
hémorragique, soit la forme bulbaire de
la sclérose latérale amyotrophique
et, dans un cas ancien, un tuberculome du tronc
cérébral. Nos observations et la
revue de la littérature permettent de
préciser que ce mouvement, associé
à l'hémiplégie, n'a pas de
latéralisation
préférentielle, qu'il peut
apparaître dès l'installation d'un
accident vasculaire à la période
de flaccidité ou, plus tard, lors de la
spasticité. Les descriptions mentionnent
soit une abduction, soit une adduction du bras.
Alors que la paralysie laisse le bras immobile
en semi-extension le long du corps du malade,
nous avons constaté un décollement
du bras du thorax, un écartement vers le
dehors (abduction) puis un mouvement de flexion
du coude ramenant la main vers le sternum
(adduction). Il est strictement contemporain du
bâillement, le bras retombant inerte
dès la fin de celui-ci. La jambe ne
présente aucun mouvement
simultané. Il est totalement
involontaire, non maîtrisable. Il est
contemporain de la paralysie et tend à
disparaître en cas de
récupération motrice. Nous pensons
que les observations de tous les auteurs
décrivent un mouvement similaire
même si les descriptions de ce bref
évènement, inhabituel et
surprenant, varient un peu.
Les observations rapportées depuis
1988 comportent l'indication de niveau de la
lésion responsable de la paralysie. Deux
localisations paraissent pouvoir engendrer ce
tableau clinique : soit existe une lésion
dans le territoire de l'artère
cérébrale moyenne, en particulier
des branches lenticulo-striées,
responsable d'un infarctus de la capsule interne
et du noyau lenticulaire (paléo-striatum)
ou du noyau caudé (néo-striatum);
soit existe une lésion bulbo-pontique,
décrite par M M
Bertolloti (1905) au cours d'un syndrome de
Millard-Gübler par tuberculome chez une
enfant de 14 ans, décrite par E.
Louwerse (1998), dans une série de
cas de Maladie de Charcot bulbaire,
décrite par R.
Töpper (2003), lors d'une thrombose
basilaire responsable d'un hémi-infarctus
du pont.
Hémiplégies et mouvements
associés
Les neurologues de la fin du XIXe
siècle parlaient déjà des
mouvements associés à
l'hémiplégie. Vulpian (1866) a
donné le nom de syncinésies
à des « mouvements qui s'effectuent
dans une partie du corps d'une façon
involontaire, au moment où ont lieu des
mouvements volontaires ou réflexes dans
une autre partie ». Lors des syndromes
pyramidaux, en général, et des
hémiplégies en particulier, les
syncinésies réalisent des
mouvements involontaires de muscles
paralysés survenant à l'occasion
de mouvements volontaires de muscles sains.
On distingue classiquement : les
syncinésies globales qui ne sont qu'une
exagération de la contracture du
côté paralysé survenant lors
d'un effort du côté sain, les
syncinésies de coordination : la
contraction volontaire d'un muscle sain
entraîne la contraction involontaire des
muscles synergiques paralysés, les
syncinésies d'imitation : un mouvement
volontaire du côté sain
déclenche une ébauche involontaire
du même mouvement du côté
paralysé.
De
Buck (1899) décrit, lui, des
mouvements associés à
l'hémiplégie : « Fait
étonnant, ils s'étendent
même aux extrêmités d'organes
qui échappent à l'empire de la
volonté. Ces processus n'accompagnent
même pas seulement les mouvements
volontaires, mais aussi les mouvements
réflexes, bâillement,
éternuement etc... ». Pierre-Marie
et A. Souques (1907) traitant de
l'hémiplégie dans la Pratique
Médico-Chirurgicale de Brissaud, Pinard
et Reclus reprend cette distinction : à
côté des syncinésies qui
n'apparaîssent qu'à la phase
spastique de l'évolution, « les
mouvements réflexes et associés
surviennent à propos d'une toux, d'un
bâillement, et se passent surtout au
membre supérieur » et peuvent
s'observer dès la phase initiale
flaccide. Bertolotti de Turin (1905) fait la
même distinction. E.
Brissaud écrit dans sa
leçon de 1893 du 1er décembre
1893: « Les bâillements spasmodiques
s'observent aussi avec une certaine
fréquence chez les
hémiplégiques. Ce n'est pas
seulement un symptôme surajouté aux
éléments spasmodiques de
l'épilepsie (Féré)
ou de l'hystérie (Charcot,
Gilles de la Tourette,
Guinon et Huet), il semble bien relever
lui-même de lésions organiques
». Nous partageons ce point de vue des
disciples de JM. Charcot bâtissant,
à l'époque, la sémiologie
neurologique clinique, et n'acceptons pas le
terme de syncinésie pour cette
élévation involontaire du bras
lors d'un bâillement.
L'éthologie décrit, chez les
vertébrés, la pandiculation ou
Rekel-syndrom comme une association d'un
bâillement avec un étirement du
tronc et des quatres membres (Selbach
et Selbach, 1953). Bertolotti (1905) et Blin
(1994), rapportant chacun une observation
similaire à celles décrites
ci-dessus, utilisent la nomination pandiculation
ou hémipandiculation. Ni la description
du mouvement involontaire du bras qui n'est pas
un étirement, ni l'absence de mouvement
d'étirement de la jambe du même
côté, ne permettent d'agréer
ce terme.
De
Buck (1899) écrit : « la
parakinésie différerait de la
synkinésie par ce fait que dans la
dernière, le rapport entre l'idée
et l'acte est parfaitement conservé et
qu'il ne s'agit que d'une simple diffusion de
l'influx nerveux, au niveau des centres moteurs,
provenant de ce que celui-ci doit être
beaucoup plus énergique pour obtenir un
effet utile, tandis que dans la première
le rapport entre l'idée et le mouvement
serait troublé ».
Il est donc
proposé le terme parakinésie
brachiale oscitante. Une parakinésie est
un mouvement anormal qui parasite, caricature ou
remplace un mouvement normal, en l'occurence
touchant le bras (brachiale). Oscitant est
emprunté du latin oscitantis, qui
bâille, comme les fièvres,
baptisées autrefois,
d'oscitantes.
Le bâillement
Le bâillement est un comportement
phylogénétiquement ancien,
d'allure réflexe, très
stéréotypé, présent
chez les vertèbrés
poïkilothermes et homéothermes, des
reptiles au cerveau rudimentaire, dit
archaïque, jusqu'aux primates humains, dans
le monde marin, aérien et terrestre. La
littérature éthologique,
neurophysiologique et neuropsychologique
décrit le bâillement associé
aux alternances des rythmes veille - sommeil,
à l'alimentation, à la
sexualité, où il apparaît
comme un comportement témoin des
mécanismes concourant à la
stimulation de la vigilance et du rôle
central de l'hypothalamus dans
l'homéostasie (H. Aubin 1988, G. Daquin
2001, O. Walusinski 2004).
Reconnaissable à l'échographie
dès quatorze
semaines de grossesse, il témoigne,
comme l'apparition de mouvements
oro-mandibulaires et de déglutition, de
la maturation fonctionnelle du tronc
cérébral et des noyaux gris
centraux alors que l'extension du cortex
fronto-pariéto-temporal se poursuit
jusqu'à 24 semaines (V. Abadie 1999).
Aucune structure cérébrale n'a
jamais pû être identifiée
comme centre du bâillement. Des arguments
cliniques et pharmacologiques indiquent que
l'hypothalamus et en particulier le noyau
paraventriculaire, les régions bulbaires
et pontiques, avec des connexions frontales chez
les primates humains et à la moelle
cervicale sont indispensables à sa
réalisation. Les muscles qui se
contractent pendant le bâillement
dépendent des nerfs crâniens V,
VII, IX, X, XI, XII, des nerfs cervicaux C1-C4
(nerf phrénique) et des nerfs dorsaux
innervant les intercostaux, muscles
respiratoires accessoires (C. Chouard
1990).
R. Provine a montré, en 1987, que le
bâillement ne modifiait en rien
l'oxygènation artérielle. Le
paradigme ancien qui, d'Hippocrate au milieu du
XX° siècle, a fait de ce
comportement un réflexe
d'oxygènation cérébrale
doit être oublié au profit d'un
paradigme neuromusculaire à commande sous
corticale.
Le noyau paraventriculaire de l'hypothalamus
contient les corps cellulaires des neurones
oxytocynergiques projettant sur l'hippocampe, la
réticulée du tronc
cérébal, le locus coeruleus. Quand
ces neurones sont stimulés par la
dopamine, les acides aminés excitateurs,
ou l'oxytocine elle-même, ils
déclenchent le comportement de
bâiller en libérant de l'oxytocine
dans ces différentes structures
sous-corticales. Cette activation
oxytocynergique est inhibée par les
opioïdes qui empêchent le
bâillement. L'activation ou l'inhibition
de ces neurones oxytocynergiques sont
corrèlées à
l'activité ou non de l'oxyde nitrique
synthétase paraventriculaire. D'autres
systèmes neuronaux sont impliqués
à titre de modulateurs. La
sérotonine, les estrogènes, la
testostérone, l'hypocrétine, la
gréline, la leptine interviennent soit au
niveau du noyau paraventriculaire soit au niveau
des noyaux moteurs du tronc
cérébral et de la moelle, voie
finale exécutive dépendante de
l'acétylcholine (A. Argiolas 1998, O.
Blin 1996, G. Daquin 2001, Sato-Suzuki I
2002).
Essai d'explications
physio-pathologiques
La libération de l'activité
des structures sous-corticales par la
levée de l'inhibition corticale est
proposée, classiquement, pour expliquer
certaines activités automatiques ou
réflexes, expérimentalement chez
l'animal décortiqué ou chez
l'homme, lors d'accidents vasculaires
cérébraux. C'est le cas pour la
myoclonie ou nystagmus du voile du palais
(Lapresle, 1984) où une destruction du
faisceau central de la calotte permet une
activité non contôlée de
l'olive bulbaire, responsable de
l'activité rythmique du voile,
réémergence fonctionnelle de
l'origine phylogénétiquement
branchiale de cette structure.
C'est le cas pour le syndrome de Foix,
Chavany, Marie (1926) ou syndrome
bi-operculaire, où persiste une
activité automatique d'expression des
émotions, de la déglutition et du
bâillement alors que la paralysie
labio-linguo-pharyngo-masticatrice est totale.
L'activité automatique dépendant
du tronc cérébral persiste alors
que le contrôle volontaire du cortex
prérolandique, de l'aire motrice
supplémentaire est interrompu par un
infarctus bi-operculaire. L'exploration par IRM
d'un cas (Ghika et Bogousslavsky, 2003), montre
cette dissociation automatico-volontaire par
disconnexion cortico-pontique. C'est, enfin, le
cas dans le "locked-in syndrom" où la
paralysie est totale, pouvant laisser seulement
une motricité occulaire volontaire
possible. Les mimiques involontaires de la
douleur, le bâillement, le cri persistent
néanmoins. La voie pyramidale
cortico-spinale est totalement interrompue au
niveau du pont. L'activité persistante
des structures extra-pyramidales,
cérébelleuses et des automatismes
médullaires archaïques expliquent la
dissociation automatico-volontaire (Bauer et al,
1980).
Un tel mécanisme de dissociation
cortico-sous-corticale peut-il expliquer la
synchronisation de l'élévation du
bras et le cycle respiratoire apparaissant lors
du bâillement par étirement du
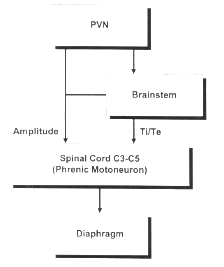
diaphragme ?
L'étude de la relation des cycles
respiratoires et de la motricité a
été initiée par Marey
(1892) grâce à son invention de la
chronophotographie, ancêtre du
cinéma. Il avait déjà
montré qu'un chien qui court, un cheval
qui galope, un gœland qui vole ont un cycle
respiratoire par foulée ou battement
d'ailes. Cette synchronisation permet une
adaptation optimale de l'oxygénation et
du travail musculaire, mais aussi, joue un
rôle important par l'effet de piston du
jeu diaphragmatique dans l'équilibre
corporel et aérodynamique. Des transferts
d'énergie entre les muscles du tronc et
les muscles respiratoires permettent une
amélioration du rendement
énergétique de la course (Bramble
et Carrier, 1983 ; Hsueh-tze, 1997 ; Viala,
1986). L'homme garde un couplage partiel avec le
balancement des membres supérieurs lors
de la marche, signe de l'activité
extra-pyramidale (Persegol et Jordan, 1991 ;
Bernasconi, 1993).
Plusieurs noyaux bulbaires et
protubérantiels concourent à
l'automatisme respiratoire. Des arguments
expérimentaux suggèrent que la
rythmogenèse ventilatoire dépend
de neurones de type pace-maker qui occupent une
région limitée appelée
complexe pré-Bötzinger, en position
ventrale par rapport au noyau ambigu et en
regard de l'émergence des racines du
douzième nerf crânien. Le rythme
engendré par les neurones pace-maker est
modulé par des réseaux
d'interneurones respiratoires responsables de
son organisation spatio-temporelle finale. Des
neurones inspiratoires, des neurones
expiratoires ont été
décrits selon la phase du cycle
ventilatoire concomitante de leur période
de décharge. Le signal rythmique
résultant des interactions neuronales est
ensuite distribué selon un schéma
spatio-temporel précis aux motoneurones
bulbaires et spinaux assurant l'innervation
motrice des muscles des voies aériennes
supérieures et des muscles respiratoires
(Bianchi et al, 1995 ; Duffin et Ezure,
1995).
Adjacent à ces noyaux, le noyau
réticulaire latéral projette vers
le cervelet, et intervient dans la coordination
sensitivo-motrice des membres, c'est la voie
spino-réticulo-cérébelleuse.
Des expériences chez le chat ont
montré qu'au cours de la décharge
des noyaux du complexe rythmique respiratoire,
ce noyau réticulaire latéral avait
lui-même une activité rythmique et
synchrone, en particulier lors de la contraction
des muscles extenseurs des membres au cours des
étirements (Arshavsky, 1978; 1986; Ezure
et Tanaka, 1997 ; Richard et Waldrop, 1989). La
bifonctionnalité des muscles intercostaux
et du diaphragme dans la posture et la
locomotion d'une part et la respiration d'autre
part (sans oublier la phonation)
nécessitent une coordination non
seulement somatomotrice mais aussi
neurovégétative, notamment
cardiovasculaire.
Schomburg (2003) a montré, chez le
chat, un contrôle
neurovégétatif sympathique
modulant l'activité cardio-locomotrice et
respiratoire avec une boucle de
rétro-contrôle
spino-protubérantielle jusqu'à
l'hypothalamus. L'adaptation cardio-respiratoire
à l'effort fait intervenir le
système neuro-végétatif, en
particulier hypothalamo-hypophysaire, en
régulant le ph sanguin, la PaCO2 mais
aussi la satiété et l'éveil
(Waldrop et al 1986 ; Yeh et al, 1997). Ces
mécanismes homéostasiques,
dépendant de structures sous corticales
anatomiquement proches, toujours
étroitement en relation, partageant leur
régulation par des neuromédiateurs
identiques, s'imbriquent avec ceux rythmant
veille - sommeil, satiété et
sexualité, c'est à dire ceux
impliqués dans le déclenchement du
bâillement (Salin-Pascual, 2001 ;
Walusinski et Deputte, 2004).
Le système moteur extra-pyramidal
exerce une influence essentielle sur les
circuits moteurs spinaux, du tronc
cérébral et du cervelet ainsi que
sur le cortex moteur lui-même. Ses
neurones cheminent du cortex
cérébral au cervelet, formant le
tractus cortico-ponto-cérébelleux
(Néo-cerebellum, développé
phylogénétiquement conjointement
avec la bipédie). C'est à travers
cette voie que le cervelet reçoit une
copie de toutes les impulsions motrices issues
du cortex cérébral (N.
Schweighofer 1998). Cette voie cortico-pontique
est disposée dans la capsule interne de
chaque côté de la voie pyramidale.
C'est à ce niveau que siège, le
plus fréquemment, la lésion
responsable de la parakinésie brachiale
oscitante, par lésion du premier neurone,
provoquant une interruption des voies
cortico-spinales et cortico-nucléaires,
mais, éventuellement aussi, des voies
extra-pyramidales cortico-striées,
cortico-rubriques, cortico-nigrales, et
cortico-réticulaires. Au niveau du pont,
les fibres font synapses dans les noyaux
pontiques dont les axones forment le second
neurone vers le cervelet (pédoncule
cérébelleux moyen). L'observation
de M. Bertolotti (1905), celles de L.
Louwerse(1998), et une de celle de R.
Töpper (2003) montrent qu'une lésion
à ce niveau peut aussi engendrer la
parakinésie brachiale oscitante.
Le paléocervelet reçoit, lui,
les influx de la mœlle
épinière par les voies
spino-cérébelleuse dorsale et
ventrale. Les deux faisceaux transmettent des
impulsions proprioceptives de la
périphérie, c'est à dire
des fuseaux neuro-musculaires et des organes
tendineux de Golgi des muscles des membres, du
tronc, et du diaphragme. Ses efférences,
après relais dans les noyaux emboliformes
et le noyau fastigial projettent vers le noyau
rouge par le pédoncule
cérébelleux supérieur. Le
paléocervelet contrôle ainsi les
muscles qui s'opposent à la gravitation,
imprime le tonus musculaire adapté
à la marche, concourt à la
coordination synergique des muscles agonistes et
antagonistes qu'elle nécessite, tout en
coordonnant le contrôle moteur du
diaphragme. Il envoie, en retour, des influx
ascendants qui se projettent sur le noyau
centro-médian du thalamus, puis vers le
noyau caudé et le putamen et assurent
ainsi une connexion avec le système
extra-pyramidal (P. Duss 1998).
L'étirement
généralisé du tronc, des
membres et du diaphragme ou pandiculation
nécessite, comme la marche normale,
l'activité fonctionnelle, sans
disconnexion, des voies
cortico-néocérébello-spinale
et
spino-archéocérébello-thalamique
couplant les commandes pyramidales et
extra-pyramidales de l'ensemble de la
musculature.
La section, quelqu'en soit
l'étiologie, des voies cortico-spinales
est responsable de la paralysie du bras pour sa
motricité volontaire. Dans certains cas,
la voie
cortico-néocérébello-spinale
est également coupée. Par contre,
la boucle proprioceptive conduisant l'influx
entre la corne médullaire
antérieure motrice, le
paléocervelet et le noyau
réticulaire latéral (voie
spino-cérébelleuse ventrale) reste
fonctionnelle. Lors du bâillement, la
puissante contraction des muscles respiratoires
représente un influx proprioceptif. Une
stimulation antidromique depuis les noyaux
respiratoires vers la corne antérieure de
la mœlle a été
démontrée expérimentalement
chez le chat (K. Ezure 1997). Une
spéculation hardie propose d'expliquer la
parakinésie brachiale oscitante comme un
mouvement du bras (semblable à celui du
balancement simultané de la respiration
lors de la marche) secondaire à un influx
moteur dans la corne antérieure
médullaire de C4 à C8, ayant pour
origine le noyau réticulaire
latéral au travers des voies
extra-pyramidales de l'archéocervelet (T.
Waldorp 1986).
Ce mécanisme explicatif est
cohérent avec l'absence de mouvement de
la jambe (section corticospinale). Il est
cohérent avec sa possibilité de
survenir tant à la phase de
flaccidité que de spasticité. La
persistance d'une voie
cortico-néocérébelleuse
intacte empêcherait son apparition. La
condition nécessaire à
l'apparition d'une parakinésie brachiale
oscitante serait donc la disjonction
cortico-nucléaire, cortico-spinale et
cortico-néocérébelleuse,
alors que la voie
spino-archéocérébelleuse
resterait fonctionnelle (D. Rosenbaum 1991). En
admettant que le bâillement est
l'extériorisation d'un comportement,
témoin d'un mécanisme
homéostasique des systèmes de
vigilance au niveau de l'hypothalamus, les
bâillements répétés,
fréquemment remarqués lors des
accidents vasculaires cérébraux,
évoqueraient une stimulation des
mécanismes de l'éveil. Lorsqu'une
lésion du système extra-pyramidal
empêche l'activité modulatrice de
la voie
cortico-néocérébelleuse,
les structures phylogénétiquement
plus anciennes permettraient de recouvrer une
motricité du bras lors de l'ampliation
maximum du diaphragme telle qu'elle se produit
au cours du bâillement.
Le système nerveux central des
vertébrés répond à
un plan d'organisation commun, et montre une
complication graduelle en rapport avec des
niveaux de vie de plus en plus
indépendants et fonctionnellement de plus
en plus élevés. Le
neuro-psychiatre américain P. MacLean
(1985) a proposé un schéma
d'organisation fonctionnelle du système
nerveux basé sur l'étude de sa
phylogenèse. Sont ainsi superposés
un cerveau ancestral « reptilien »
(tronc cérébral et noyaux gris
centraux), lieu d'origine du bâillement,
un cerveau « paléomammalien »
(système limbique) commun à tous
les mammifères, interface synaptique et
humorale, siège du bâillement
d'émotivité des singes, un cerveau
« néomammalien »
caractérisé par le
développement cortical chez l'homme, en
particulier des lobes frontaux siège de
la « contagion » du bâillement
(O.Walusinski 2004). La dissection de ces
étages fonctionnels, dans certaines
localisations d'accidents vasculaires
cérébraux, laisse
réapparaître des
fonctionnalités normalement
inhibées par une structure
phylogénétiquement plus
récente et fonctionnellement plus
élaborée. La pathologie humaine
permet ainsi d'appréhender que la
coordination de la régulation de la
température corporelle, de la
respiration, de la locomotion et de
l'éveil subit un perfectionnement, une
adaptation graduellement de plus en plus
complexe et précise, depuis les reptiles
jusqu'aux primates.
Argiolas
A, Melis M.R. The neuropharmacology of
yawning Eur Pharmacol; 1998; 343; 1; 1-16
Arshavsky
YI. et al Messages convey by
spino-cerebellar pathways during scratching in
the cat Brain Res 1978; 151; 479-506
Arshavsky YI. et al Cerbellum and rhythmical
movements Springer Berlin 1986
Aubin H.J, Garma
L Le bâillement Psychiatrie et
Psychobiologie; 1988; 3; 275-286
Bauer G. et al
Involuntary motor phenomena in the locked in
syndrome J Neurology 1980; 223; 191-198
Bernasconi
P, Kohl J. Analysis of coordination between
breathing and exercise rhythms in man J
Physiology 1993; 471; 693-706
Bertolotti
M. Etude sur la pandiculation automatique
des hémiplégiques Rev Neurol 1905;
2; 19; 953-959
Bianchi
AL, Denavit-Saubié M, Champagnat J.
Central control of breathing in mammals :
neuronal circuitry, membrane properties, and
neurotransmitters. Physiological Reviews 1995;
75; 1-45
Blin
O. Le bâillement en
neuropsychopharmacologie clinique Lettre du
pharmacologue 1996; 10; 217-219
de
Buck Classification des mouvements anormaux
associés à
l'hémiplégie Rev Neurol 1899; 6;
361-374
Duan
Y et al Cardiorespiratory components of
defense reaction elicited from paraventricular
nucleus Physiol Behav 1997; 61; 2; 325-330
Ezure K,
Tanaka I. Convergence of central respiratory
and locomotor rhythms onto sigle neurons of the
lateral reticular nucleus Exp Brain Res 1997;
113; 230-242
Foix C, Chavany
JA, Marie J. Diplégie
facio-linguo-masticatrice d'origine
cortico-souscorticale sans paralysie des membres
Rev Neurol 1926, 33, 214-219
Fugl-Meyer
AR,, Linderholm H et al. Restrictive
ventilatory dysfunction in stroke: its relation
to locomotor function Scand J Rehabil Med 1983;
Suppl 9; 118-124
Furtado D.
Provocation spinale d'un réflexe de
bâillement. Rev d'oto neuro
ophtalmologie.1951;23(1):
Ghika
J, Bogousslavsky J. Dissociated preservation
of automatic-voluntary jaw movements in a
patient with biopercular and unilateral pontine
infarcts Eur Neurol 2003, 50, 185-188
Heusner A P
Yawning and associated phenomena Physiological
Review 1946; 25; 156-168
Hsueh-tze
L. Mechanical links between locomotion and
beathing: can you breath with your legs ? News
Physiol. Sci. 1997; 12; 273-278 télécharger
PDF
Lanari A,
Delbono O The yawning and stretching sign in
hemiplegics Medicina (B Aires) 1983; 43; 3;
355-356
Lapresles
J La voie dento-olivaire: sa mise en
évidence, son trajet, sa signification
Bull Acd Nat Med 1984, 168, 3-, 336-341
Liecey Nouvelle
observation de bâillement convulsif
périodique Le Courrier Médical
1879; 29; 334-336
Liégey Deux
observations de bâillements intermittents
Gazette médicale de Strasbourg 1851;
p118-119
Louwerse E
Forced yawning as a pseudobulbar sign in
amyotrophic lateral sclerosis J Neuroscience
Research 1998, sup, 392
MacLean P
Evolutionay psychiatry and the triune brain;
Psychological Medecine 1985; 15; 219-221
Marey
EJ La photographie du mouvement G.
Carré ed Paris 1892, 97p (le
musée)
Mignot E., et al Sleeping with the
hypothalamus Nature Neuroscience 2002; 5;
1071-1075
Morin
D, Viala D Coordinations of locomotor and
respiratory rhythms in vitro are critically
dependent on hindlimb sensory inputs. J Neurosci
2002; 22; 11; 4756-65. télécharger
PDF
Mulley G
Assoctiated reactions in the hemiplegic arm
Scand J Rehab Med 1982; 14; 117-120
Ogle JW
Arm rising during yawning The Medical Times and
Gazette. 28 fébruary 1863 p 213
Persegol
L, Jordan M et al. Evidence for central
entrainment of the medullary respiratory pattern
by the locomotor pattern in the rabbit. Exp
Brain Res 1988; 71; 1; 153-62
Persegol
L, Jordan M et al. Evidence for the
entrainment of breathing by locomotor pattern in
human. J Physiol (Paris) 1991; 85; 1; 38-43
Pierre
Marie La Pratique Neurologique
Hémiplégie, mouvements
associés Masson 1911, p477-480
Pierre
Marie et André Léri Mouvements
involontaires dans les membres paralysés
1911 Nouveau Traité de médecine et
de thérapeutique Brouardel, Gilbert,
Thoinot JB Baillière Ed 1911
p283-291
Quoirin E.
Elévation involontaire du membre
supérieur chez
l'hémiplégique lors d'un
bâillement Thèse doctorat en
médecine Poitiers 2002
Richard
CA, Waldrop TG et al. The nucleus
reticularis gigantocellularis modulates the
cardiopulmonary responses to central and
peripheral drives related to exercise Brain Res
1989; 482; 1; 49-56
Romberg
MH A manual of the nervous diseases of man.
Berlin - London 1853
Rosenbaum D. Human Motor Control 300 p
Academic Press 1990
Salin-Pascual
R. et al Hypothalamic regulation of sleep
Neuropsychopharmacology 2001; 25; S21-S27
Sato-Suzuki I,
I Kita, Y, Seki, M Oguri, H Arita. Cortical
arousal induced by microinjection of orexins
into the paraventricular nucleus of the rat
Behavioural Brain Research 2002; 128;
169-177
Schiller
F Yawning ? J History of the Neurosciences
2002; 11; 4; 392-401
Schomburg
ED et al Rhytmic phrenic, intercostal and
sympathetic activity in relation to limb and
trunk motor activity in spinal cats Neuroscience
Research 2003: 46; 229-240
Selbach C, H
Selbach Das Räkel-Syndrom als
Wirkungsfolge eines biologischen Regelsystems.
Monatschr.f. Psychiat u Neurol; 1953; 125;
671-82.
Thomson HC
Associated movements in hemiplegia : their
origin and physiological significance Brain
1903; 26; 515-523
Töpper
R, Mull M, Nacimento W Involuntary
stretching during yawning in patients with
pyramidal tract lesions: further evidence for
the existence of an independent emotional motor
system European J Neurology 2003; 10;
495-499
Trautmann R Le
bâillement Thèse Bordeaux; 1901-02;
N° 40; 86 pages
Viala
D Evidence for direct reciprocal
interactions between the central rhythm
generators for spinal "respiratory" and
locomotor activities in the rabbit Exp Brain Res
1986; 63; 2; 225-32
Vulpian
A Leçons sur la physiologie
générale et comparée du
système nerveux faites au Museum
d'histoire naturelle. Rédigées par
E.Brémond. Paris, Baillière,
1866
Vulpian
A Maladies du système nerveux Paris,
Doin, 1879
Waldrop
TG et al Control of respiration by the
hypothalamus and feedback from contracting
muscles in cats Respir. Physiol 1986; 64;
317-328
Walusinski O,
Deputte BL Le bâillement :
phylogenèse, éthologie,
nosogénie Revue Neurologique
2004;160(11):1011-1021
Walshe FMR
On certain tonic or postural reflexes in
hemiplegia with special reference to the so
called "associated movements". Brain,
1923;46:1-37
Wimalaratna HS,
Capildeo R. Is yawning a brainstem
phenomenon ? a stroke patient who stretched his
hemiplegic arm during yawning Lancet 1988; 1;
8580; 300
Walshe FMR. On
certain tonic or postural reflexes in hemiplegia
with special reference to the so called «
associated movements » Brain 1923; 46;
1-37
Yeh E
et al The paraventricular nucleus of the
hypothalamus influences respiratory timing and
activity in the rat Neurosci Let 1997; 232;
63-66
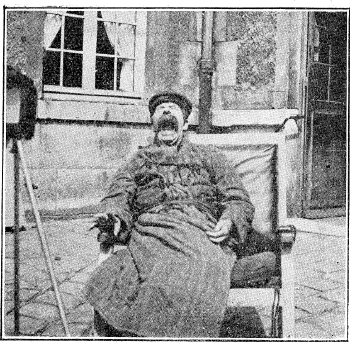
Dans la collection des clichés de
Pierre Marie en 1921, ce patient pseudo bulbaire
au rire et pleurer spasmodique, parésie
des 4 membres, bouge les doigts droits lors de
son bâillement !
The nervous system of the human body as
explained in a series of papers read before the
Royal Society of London with an appendix of
cases and consultations on nervous
diseases.
Charles Bell - London. Henry Renshaw Ed.
1844. p411
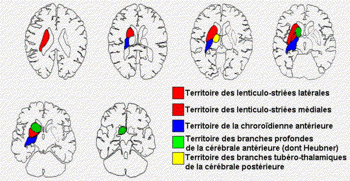
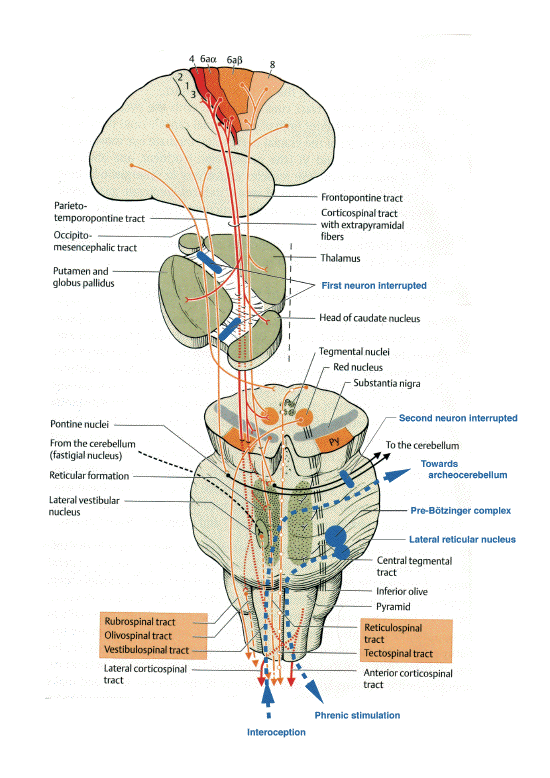
Schematic representation of the
extrapyramidal tract and the localizations of
lesions responsible for parakinesia brachialis
oscitans. (adapted from Duus' Topical Diagnosis
in Neurology, Thieme ed. 2005)